

This is a preprint.
An alternative route for β-hydroxybutyrate metabolism supports fatty acid synthesis in cancer cells
- PMID: 39554134
- PMCID: PMC11565918
- DOI: 10.1101/2024.10.31.621317
An alternative route for β-hydroxybutyrate metabolism supports fatty acid synthesis in cancer cells
Update in
-
An alternative route for β-hydroxybutyrate metabolism supports cytosolic acetyl-CoA synthesis in cancer cells.Nat Metab. 2025 Oct;7(10):2033-2044. doi: 10.1038/s42255-025-01366-y. Epub 2025 Sep 8. Nat Metab. 2025. PMID: 40921754 Free PMC article.
Abstract
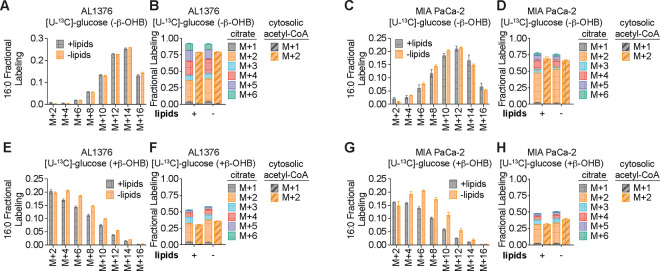
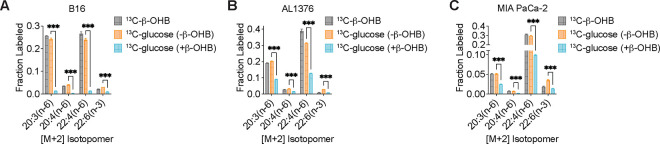
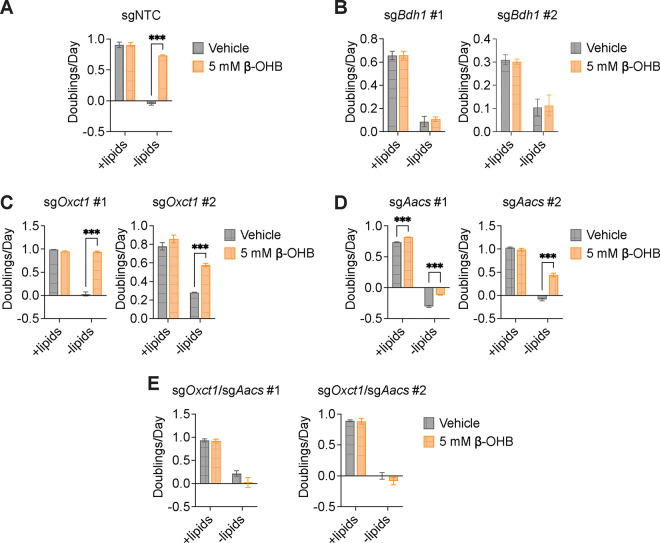
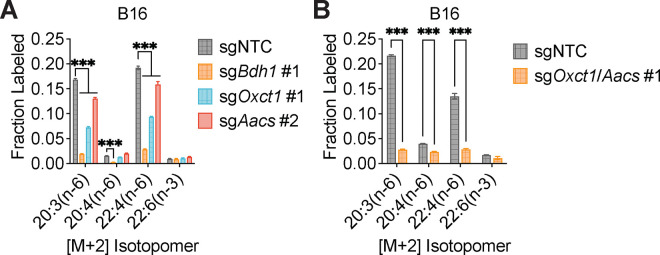
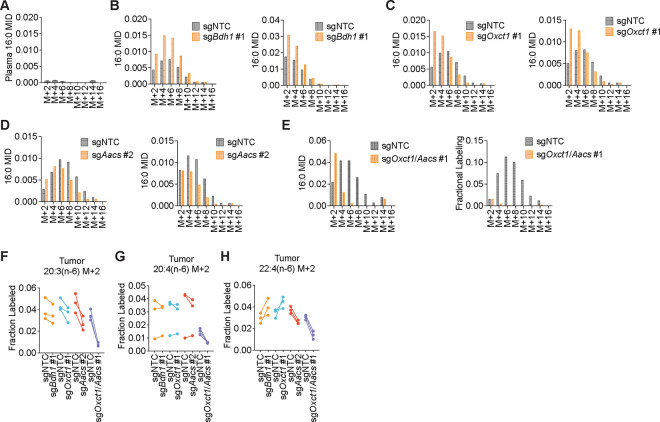
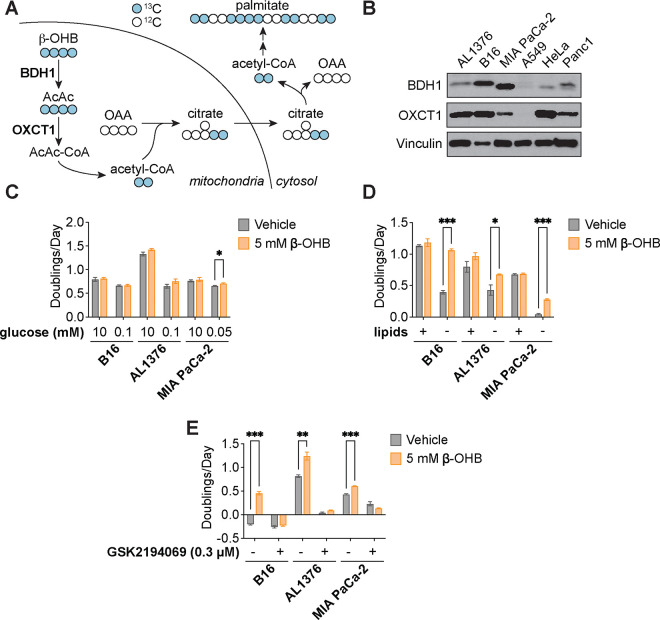
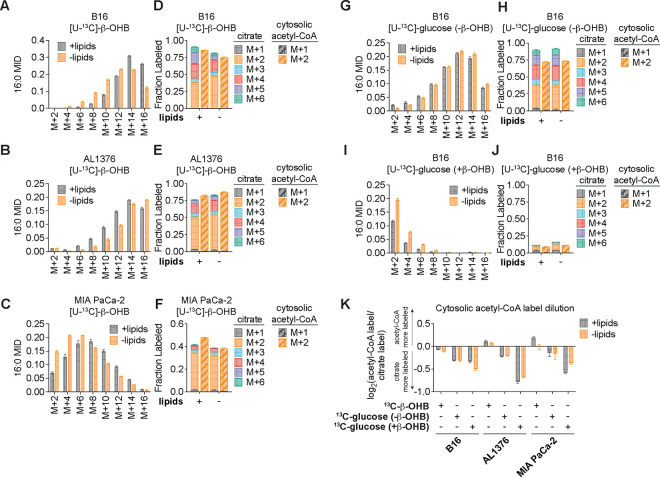
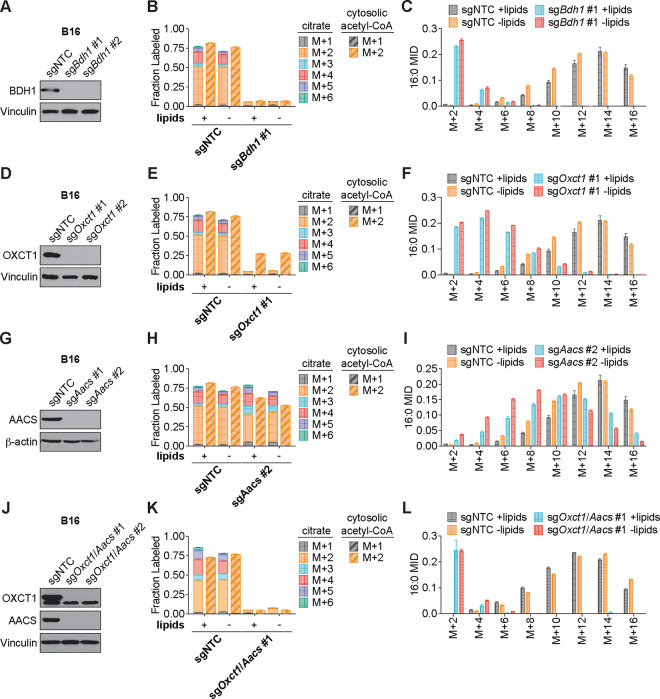
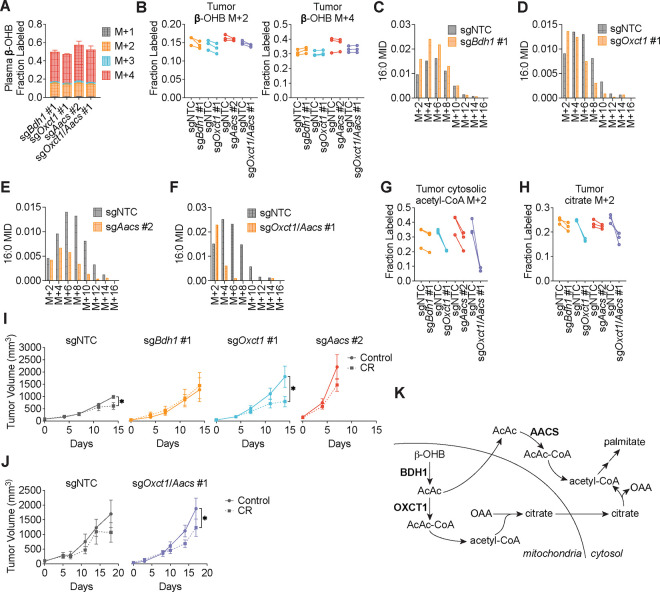
Cancer cells are exposed to diverse metabolites in the tumor microenvironment that are used to support the synthesis of nucleotides, amino acids, and lipids needed for rapid cell proliferation1-3. Recent work has shown that ketone bodies such as β-hydroxybutyrate (β-OHB), which are elevated in circulation under fasting conditions or low glycemic diets, can serve as an alternative fuel that is metabolized in the mitochondria to provide acetyl-CoA for the tricarboxylic acid (TCA) cycle in some tumors4-7. Here, we discover a non-canonical route for β-OHB metabolism, in which β-OHB can bypass the TCA cycle to generate cytosolic acetyl-CoA for de novo fatty acid synthesis in cancer cells. We show that β-OHB-derived acetoacetate in the mitochondria can be shunted into the cytosol, where acetoacetyl-CoA synthetase (AACS) and thiolase convert it into acetyl-CoA for fatty acid synthesis. This alternative metabolic routing of β-OHB allows it to avoid oxidation in the mitochondria and net contribute to anabolic biosynthetic processes. In cancer cells, β-OHB is used for fatty acid synthesis to support cell proliferation under lipid-limited conditions in vitro and contributes to tumor growth under lipid-limited conditions induced by a calorie-restricted diet in vivo. Together, these data demonstrate that β-OHB is preferentially used for fatty acid synthesis in cancer cells to support tumor growth.
Conflict of interest statement
Competing interests: R.G.J. is a scientific advisor to Servier Pharmaceuticals and is a member of the Scientific Advisory Board of Immunomet Therapeutics. All other authors declare no competing interests.
Figures
References
-
- Hanahan D. & Weinberg R. A. Hallmarks of cancer: the next generation. Cell 144, 646–74 (2011). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources