

This is a preprint.
Transcriptional profiles of murine oligodendrocyte precursor cells across the lifespan
- PMID: 39554158
- PMCID: PMC11565715
- DOI: 10.1101/2024.10.27.620502
Transcriptional profiles of murine oligodendrocyte precursor cells across the lifespan
Update in
-
Transcriptional profiles of mouse oligodendrocyte precursor cells across the lifespan.Nat Aging. 2025 Apr;5(4):675-690. doi: 10.1038/s43587-025-00840-2. Epub 2025 Mar 31. Nat Aging. 2025. PMID: 40164771
Abstract
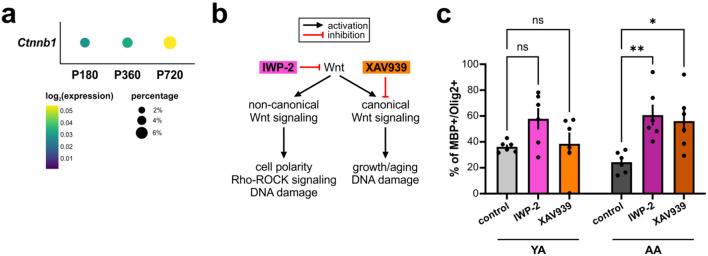
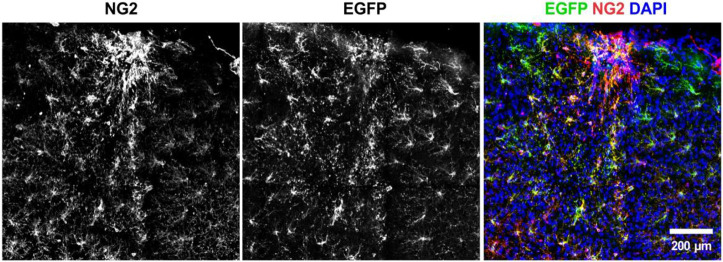
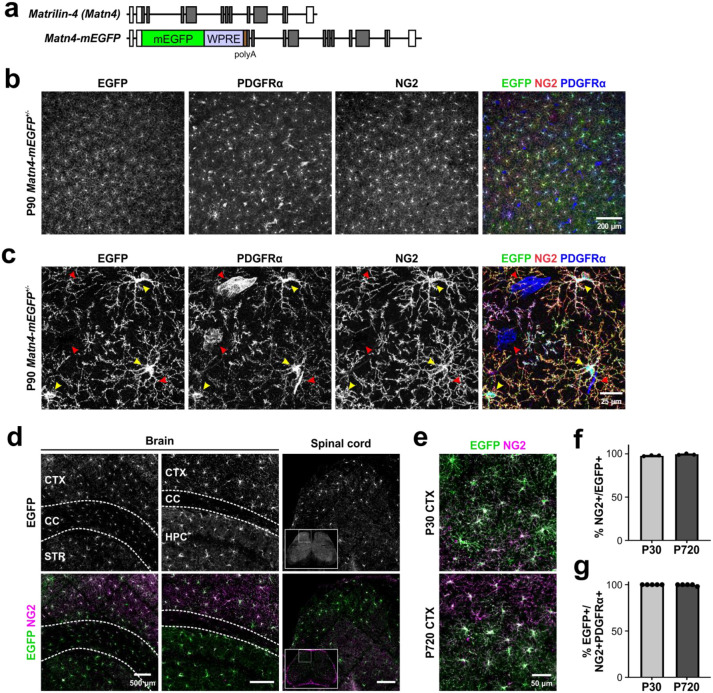
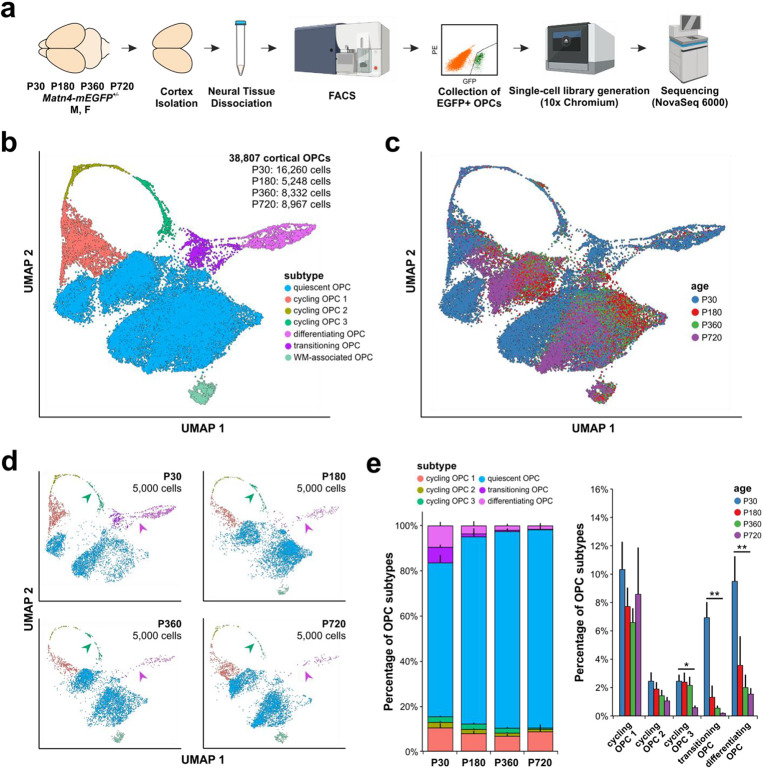
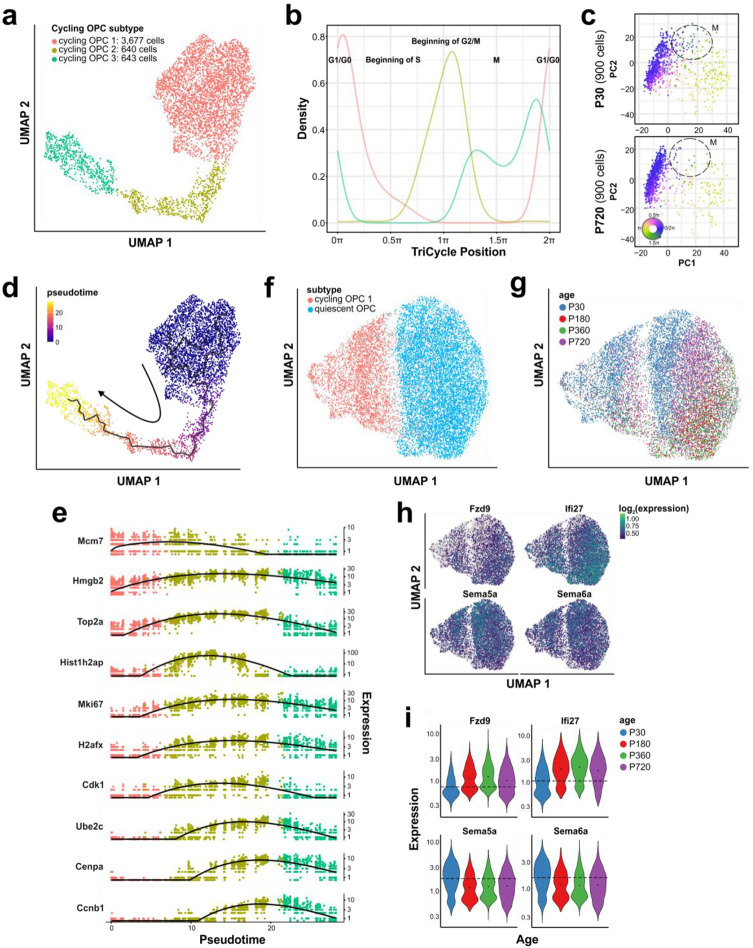
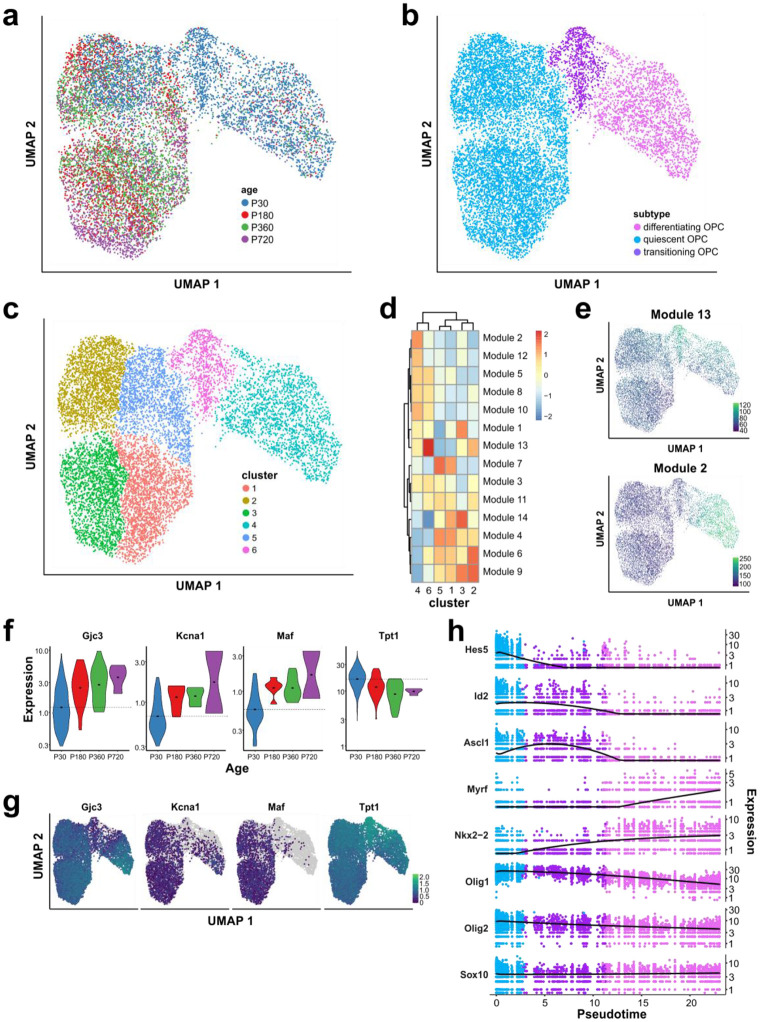
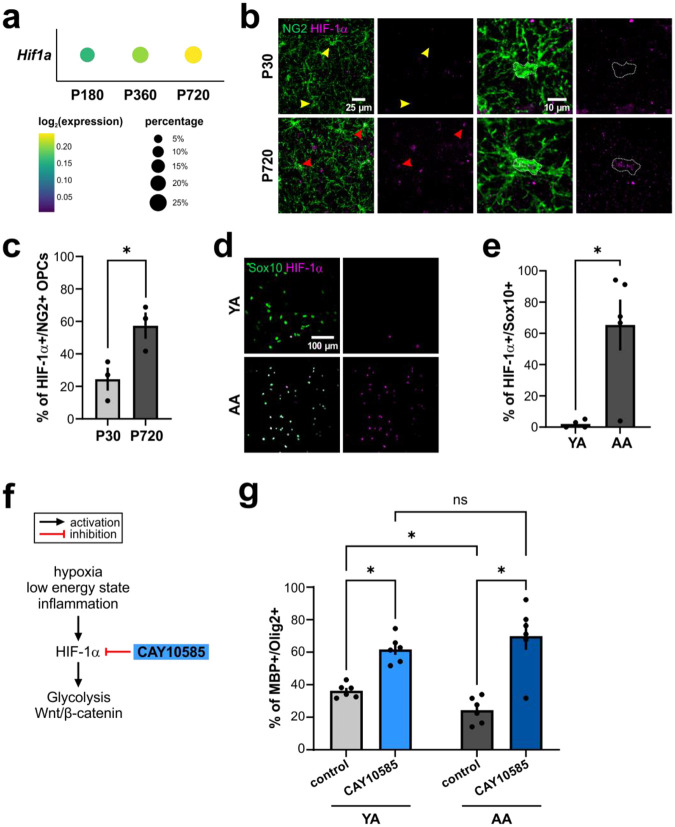
Oligodendrocyte progenitor cells (OPCs) are highly dynamic, widely distributed glial cells of the central nervous system (CNS) that are responsible for generating myelinating oligodendrocytes during development. By also generating new oligodendrocytes in the adult CNS, OPCs allow formation of new myelin sheaths in response to environmental and behavioral changes and play a crucial role in regenerating myelin following demyelination (remyelination). However, the rates of OPC proliferation and differentiation decline dramatically with aging, which may impair homeostasis, remyelination, and adaptive myelination during learning. To determine how aging influences OPCs, we generated a novel transgenic mouse line that expresses membrane-anchored EGFP under the endogenous promoter/enhancer of Matrilin-4 (Matn4-mEGFP) and performed high-throughput single-cell RNA sequencing, providing enhanced resolution of transcriptional changes during key transitions from quiescence to proliferation and differentiation across the lifespan. Comparative analysis of OPCs isolated from mice aged 30 to 720 days, revealed that aging induces distinct inflammatory transcriptomic changes in OPCs in different states, including enhanced activation of HIF-1α and Wnt pathways. Inhibition of these pathways in acutely isolated OPCs from aged animals restored their ability to differentiate, suggesting that this enhanced signaling may contribute to the decreased regenerative potential of OPCs with aging. This Matn4-mEGFP mouse line and single-cell mRNA datasets of cortical OPCs across ages help to define the molecular changes guiding their behavior in various physiological and pathological contexts.
Figures

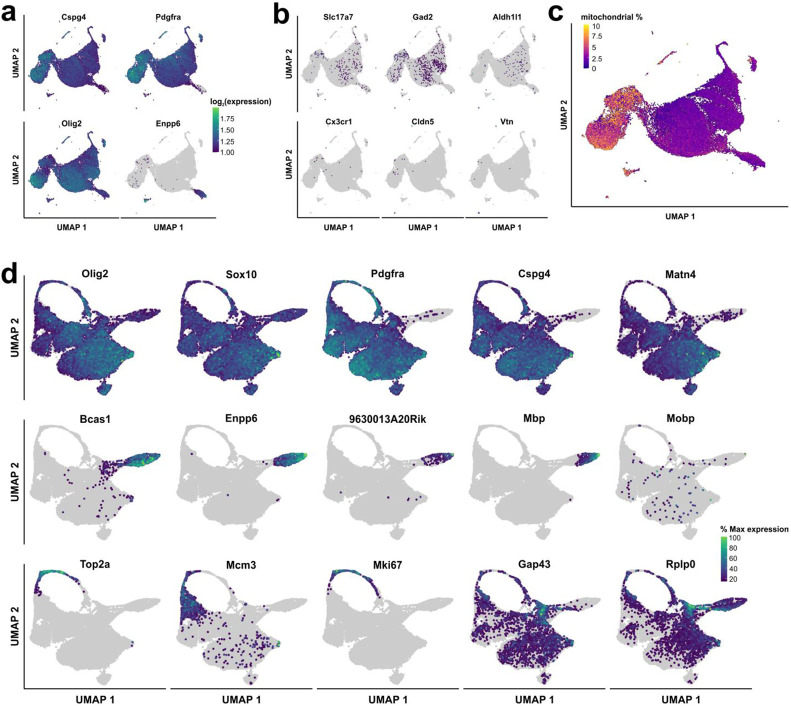

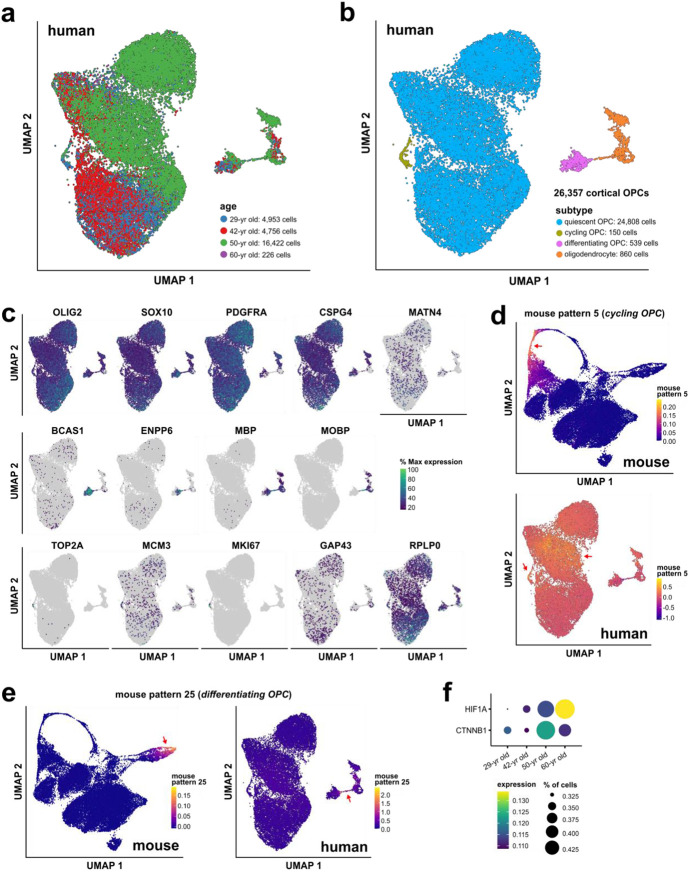
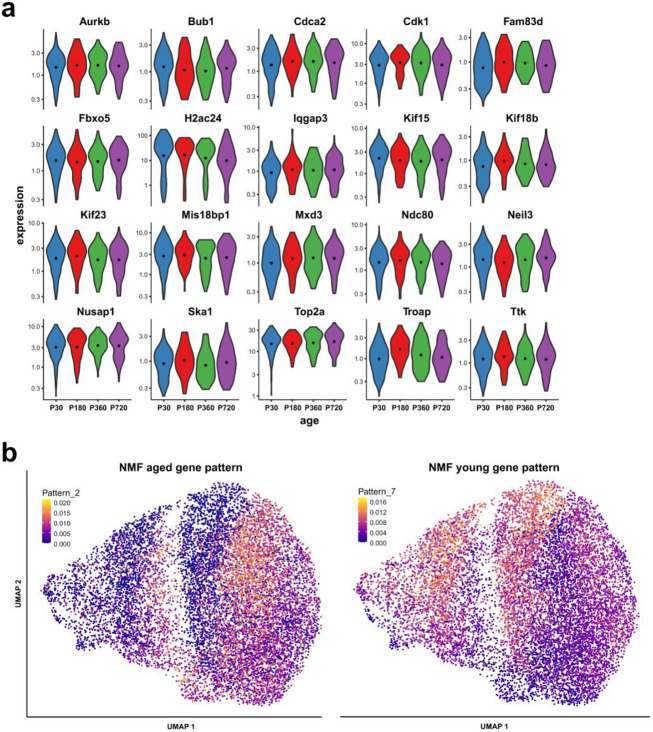
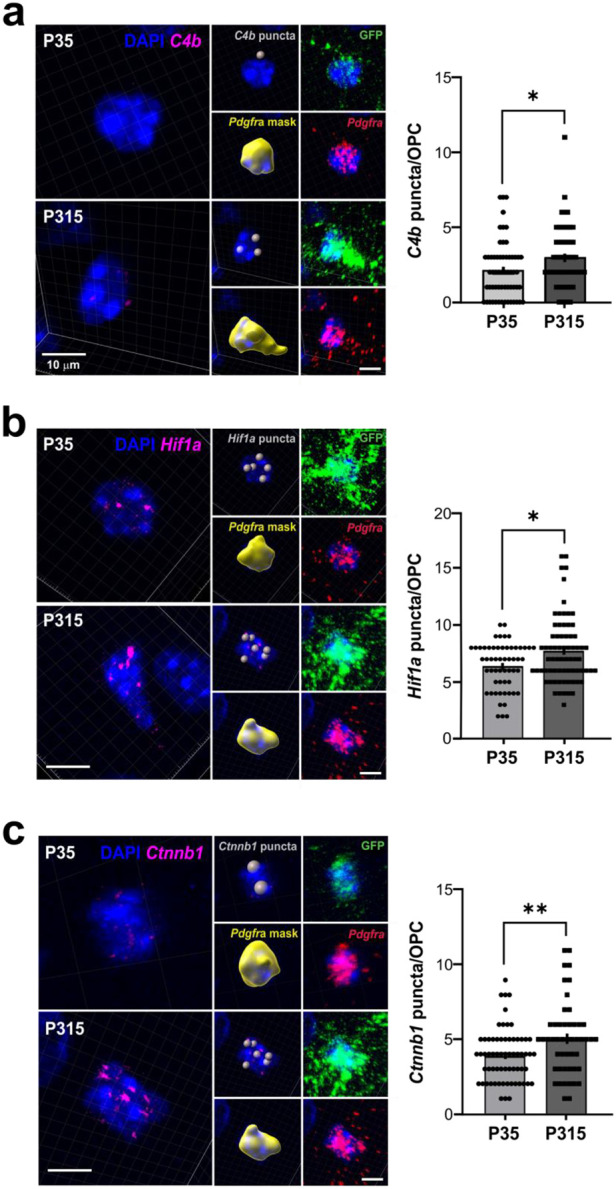

References
-
- Salas A. et al. JAK-STAT pathway targeting for the treatment of inflammatory bowel disease. Nat Rev Gastroenterol Hepatol 17, 323–337 (2020). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources