

This is a preprint.
Dynamic Kidney Organoid Microphysiological Analysis Platform
- PMID: 39554191
- PMCID: PMC11565751
- DOI: 10.1101/2024.10.27.620552
Dynamic Kidney Organoid Microphysiological Analysis Platform
Abstract
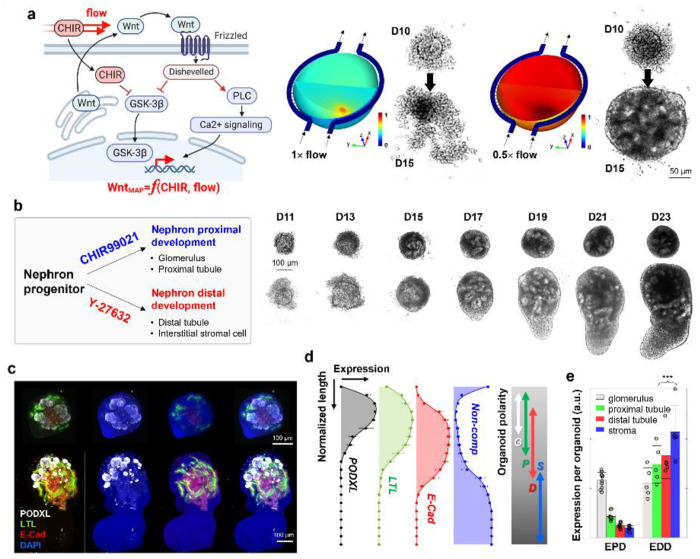
Kidney organoids, replicating human development, pathology, and drug responses, are a promising model for advancing bioscience and pharmaceutical innovation. However, reproducibility, accuracy, and quantification challenges hinder their broader utility for advanced biological and pharmaceutical applications. Herein, we present a dynamic kidney organoid microphysiological analysis platform (MAP), designed to enhance organoid modeling and assays within physiologically relevant environments, thereby expanding their utility in advancing kidney physiology and pathology research. First, precise control of the dynamic microenvironment in MAP enhances the ability to fine-tune nephrogenic intricacies, facilitating high-throughput and reproducible human kidney organoid development. Also, MAP's miniaturization of kidney organoids significantly advances pharmaceutical research by allowing for detailed analysis of entire nephron segments, which is crucial for assessing the nephrotoxicity and safety of drugs. Furthermore, the MAP's application in disease modeling faithfully recapitulates pathological development and functions as a valuable testbed for therapeutic exploration in polycystic kidney diseases. We envision the kidney organoid MAP enhancing pharmaceutical research, standardizing processes, and improving analytics, thereby elevating the quality and utility of organoids in biology, pharmacology, precision medicine, and education.
Figures



References
-
- Godin R. E., Robertson E. J. & Dudley A. T. Role of BMP family members during kidney development. International Journal of Developmental Biology 43, 405–411 (2003). - PubMed
-
- Carroll T. J. & McMAHON A. P. Secreted molecules in metanephric induction. Journal of the American Society of Nephrology 11, S116–S119 (2000). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources