

Frontotemporal network contribution to occluded face processing
- PMID: 39556727
- PMCID: PMC11621840
- DOI: 10.1073/pnas.2407457121
Frontotemporal network contribution to occluded face processing
Abstract
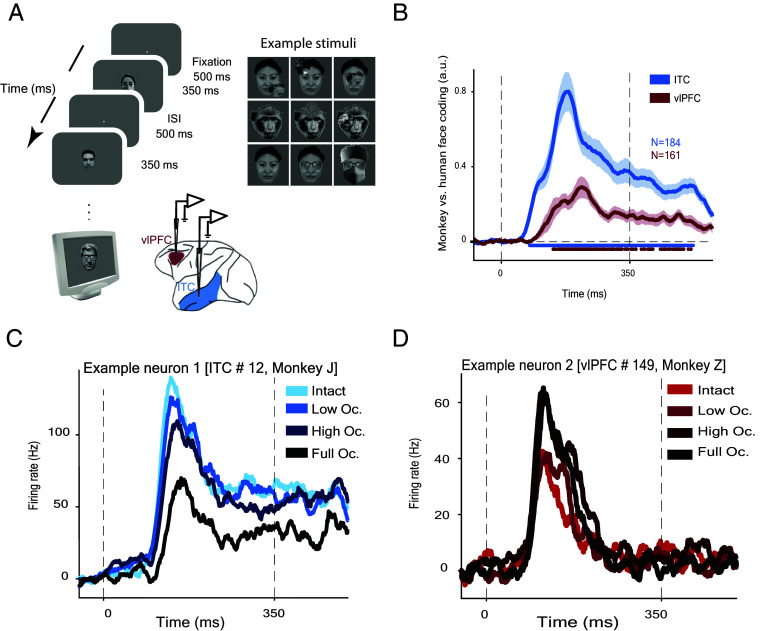
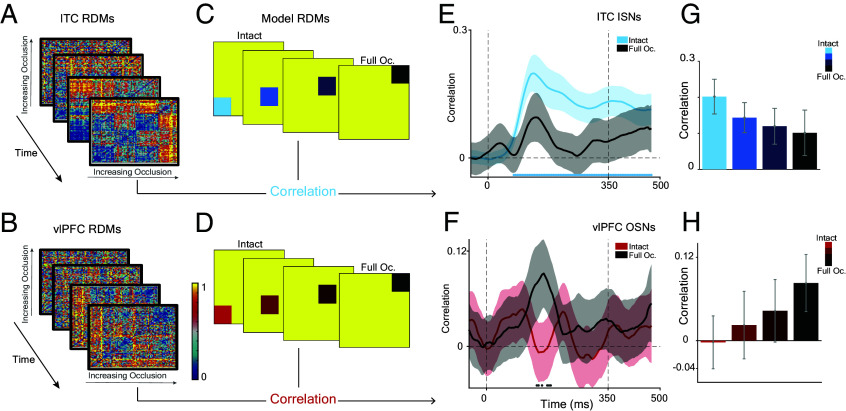
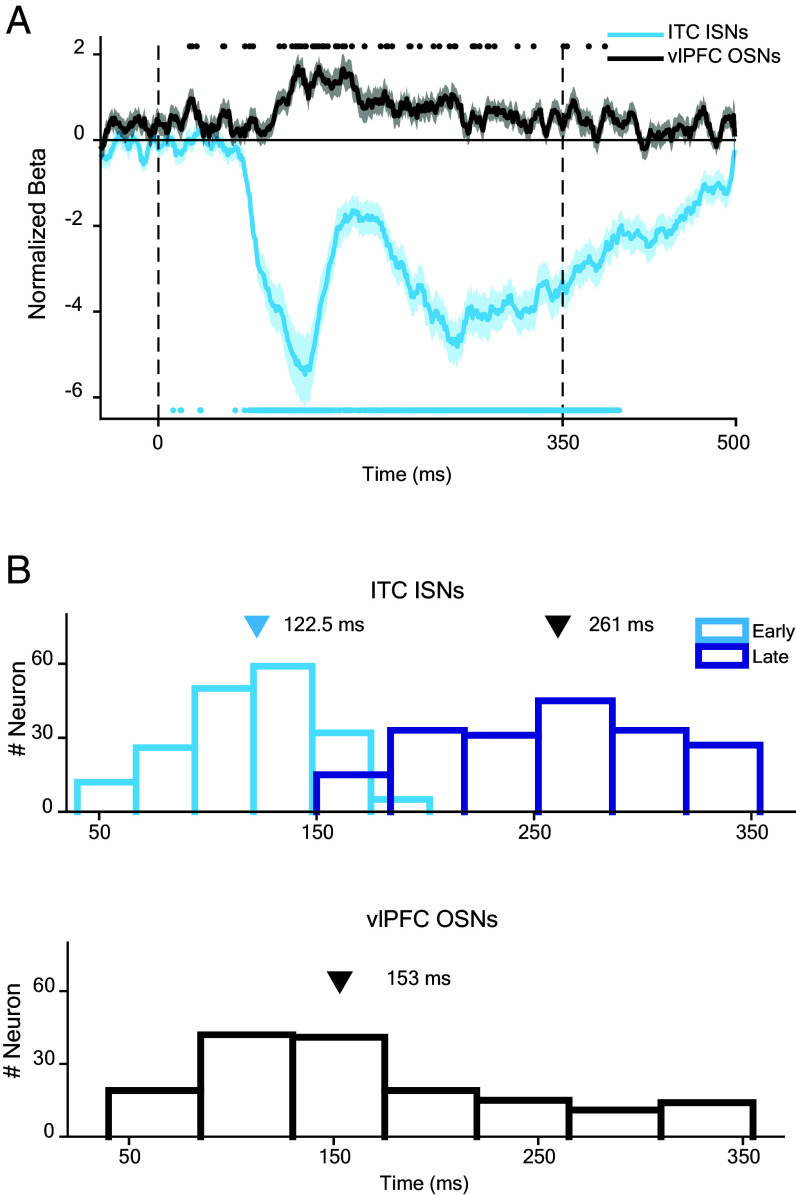
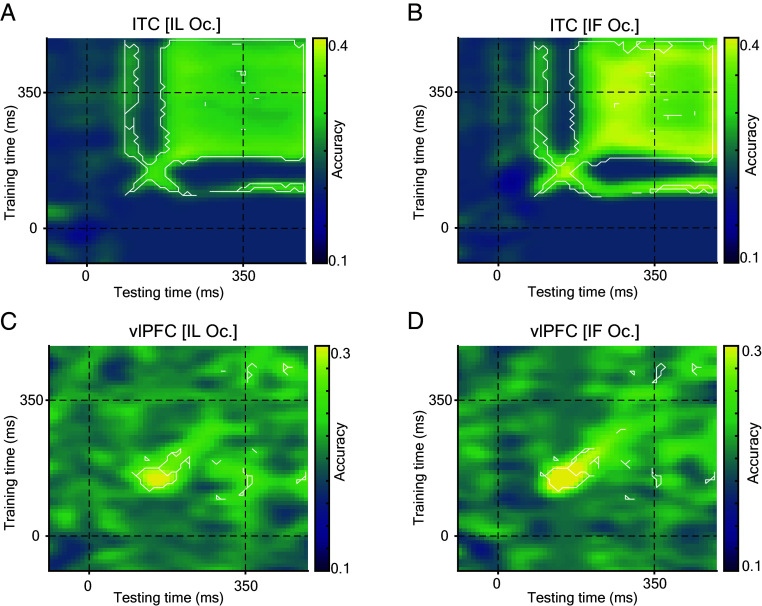
Primates are known for their exceptional ability to recognize faces. However, we still have much to learn about how their brains process faces when they are partially hidden. When we cover parts of a face, it affects how our brains respond, even though we still perceive the face as a whole. This suggests that complex brain networks are at work in understanding partially hidden faces. To explore this further, we studied two brain regions, the ventrolateral prefrontal cortex (vlPFC) and the inferior temporal cortex (ITC), while showing primate images of faces with parts occluded. We found that vlPFC neurons were more active when faces were partially covered, while ITC neurons preferred fully visible faces. Interestingly, the ITC seemed to process occluded faces in a separate phase after the vlPFC responded. Our research revealed a coordinated effort between these brain regions based on the level of facial obstruction. Specifically, the vlPFC seemed to play a crucial role, driving the representation of occluded faces in the later phase of ITC processing. Importantly, we also found that the brain processes occluded faces differently from those that are fully visible, suggesting specialized mechanisms for handling these situations. These findings highlight the importance of feedback from the vlPFC in understanding occluded faces in the ITC region of the brain. Understanding these neural processes not only enhances our understanding of how primates perceive faces but also provides insights into broader aspects of visual cognition.
Keywords: face recognition; inferior temporal cortex; occlusion; ventrolateral prefrontal cortex.
Conflict of interest statement
Competing interests statement:The authors declare no competing interest.
Figures





References
-
- Burton A. M., Jenkins R., Hancock P. J., White D., Robust representations for face recognition: The power of averages. Cogn. Psychol. 51, 256–284 (2005). - PubMed
-
- Jacques C., et al. , The neural basis of rapid unfamiliar face individuation with human intracerebral recordings. Neuroimage 221, 117174 (2020). - PubMed
-
- Taubert J., Weldon K. B., Parr L. A., Robust representations of individual faces in chimpanzees (Pan troglodytes) but not monkeys (Macaca mulatta). Anim. Cogn. 20, 321–329 (2017). - PubMed
-
- Thatere A., Meshram A., Verma P., Jirapure A., Eds., “Face recognition under occlusion: An efficient handcrafted feature & SVM based approach” in 2024 IEEE International Conference on Interdisciplinary Approaches in Technology and Management for Social Innovation (IATMSI), Thatere A., Meshram A., Verma P., Jirapure A., Eds. (IEEE, 2024).
-
- Fu M., Wang Z., Fan D., Wu H., Eds., “Occlusion face recognition based on improved attention mechanism” in International Conference on Computer Graphics, Artificial Intelligence, and Data Processing (ICCAID 2022), Fu M., Wang Z., Fan D., Wu H., Eds. (SPIE, 2023).
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
