Mechanistic basis of atypical TERT promoter mutations
- PMID: 39557834
- PMCID: PMC11574208
- DOI: 10.1038/s41467-024-54158-5
Mechanistic basis of atypical TERT promoter mutations
Abstract
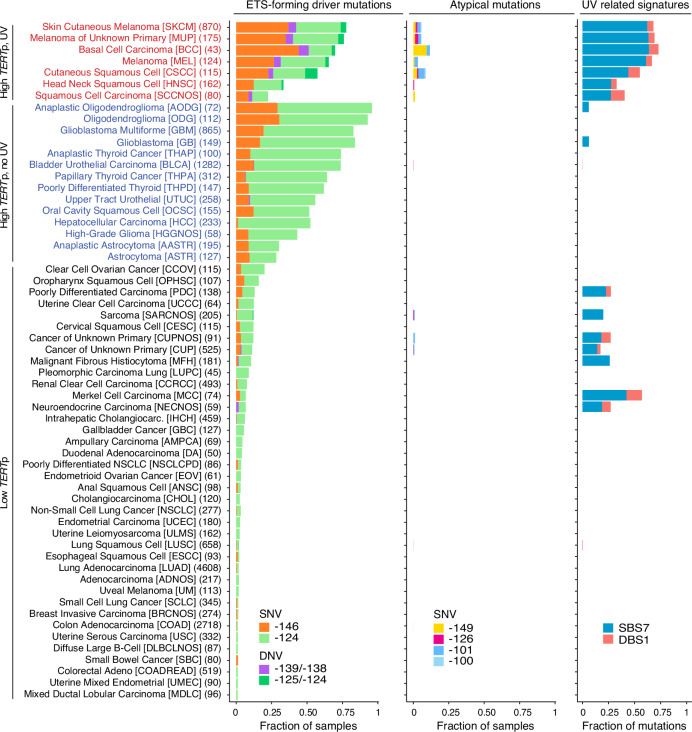
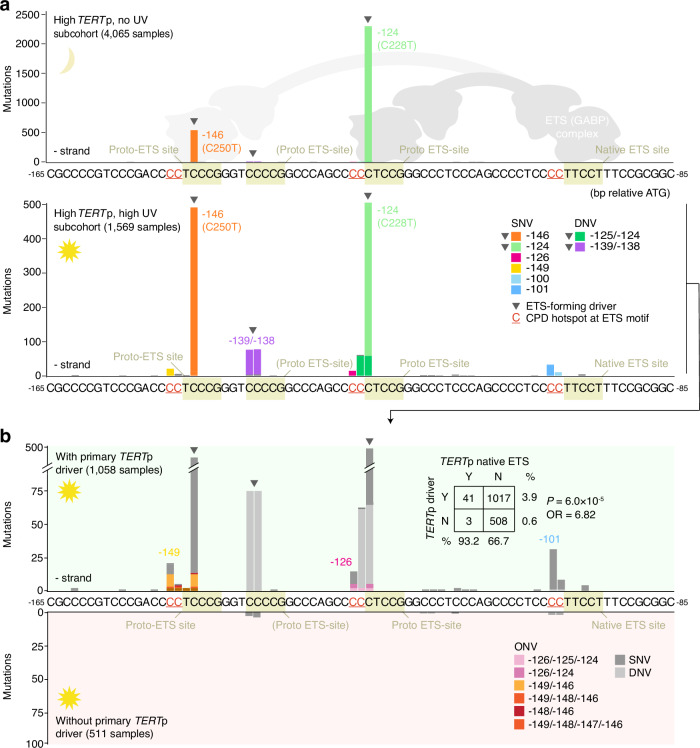
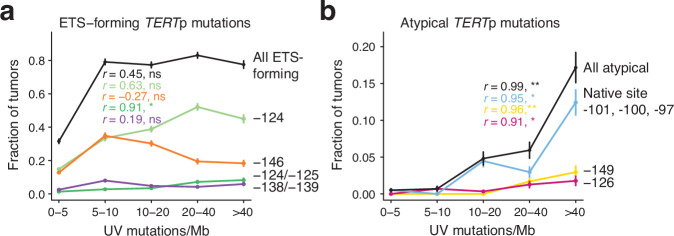
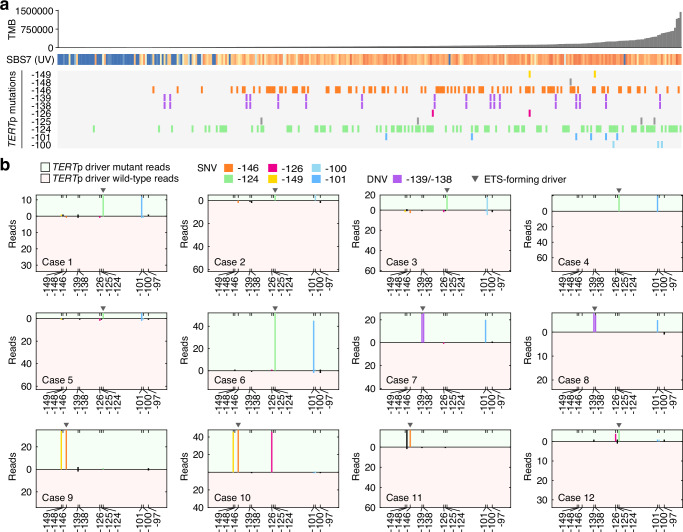
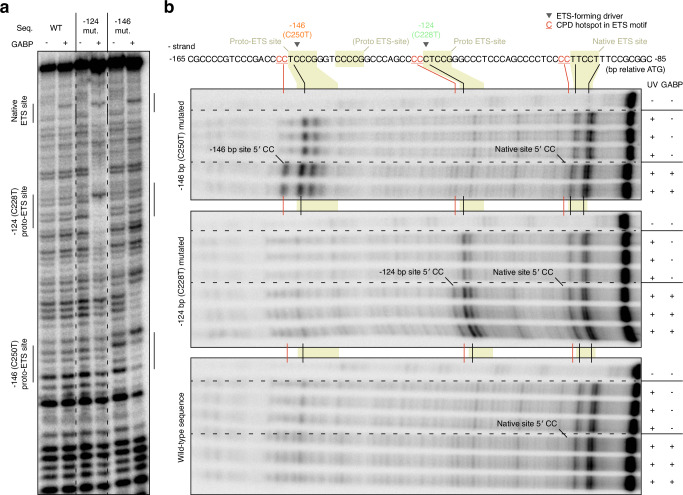
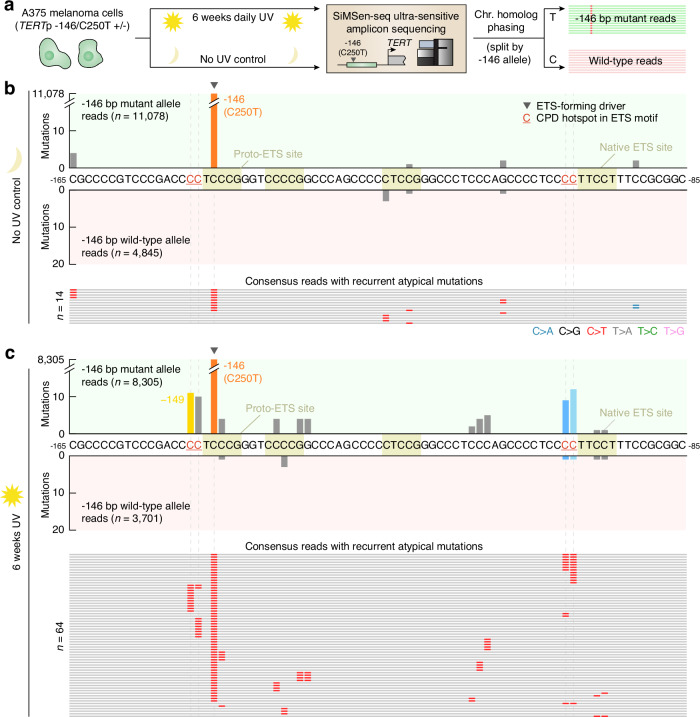
Non-coding mutations in the TERT promoter (TERTp), typically at one of two bases -124 and -146 bp upstream of the start codon, are among the most prevalent driver mutations in human cancer. Several additional recurrent TERTp mutations have been reported but their functions and origins remain largely unexplained. Here, we show that atypical TERTp mutations arise secondary to canonical TERTp mutations in a two-step process. Canonical TERTp mutations create de novo binding sites for ETS family transcription factors that induce favourable conditions for DNA damage formation by UV light, thus creating a hotspot effect but only after a first mutational hit. In agreement, atypical TERTp mutations co-occur with canonical driver mutations in large cancer cohorts and arise subclonally specifically on the TERTp driver mutant chromosome homolog of melanoma cells treated with UV light in vitro. Our study gives an in-depth view of TERTp mutations in cancer and provides a mechanistic explanation for atypical TERTp mutations.
© 2024. The Author(s).
Conflict of interest statement
Figures