

Boosting neuraminidase immunity in the presence of hemagglutinin with the next generation of influenza vaccines
- PMID: 39562599
- PMCID: PMC11577023
- DOI: 10.1038/s41541-024-01011-x
Boosting neuraminidase immunity in the presence of hemagglutinin with the next generation of influenza vaccines
Abstract
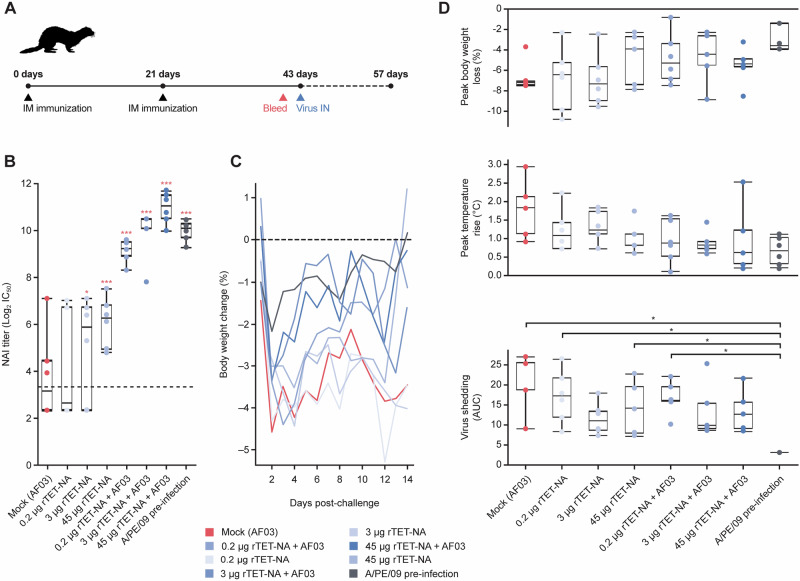
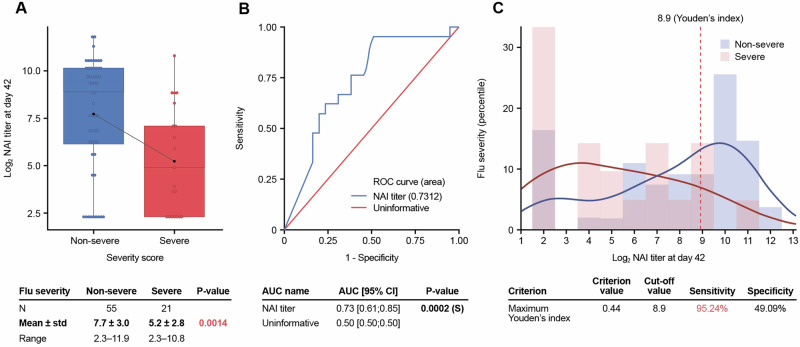
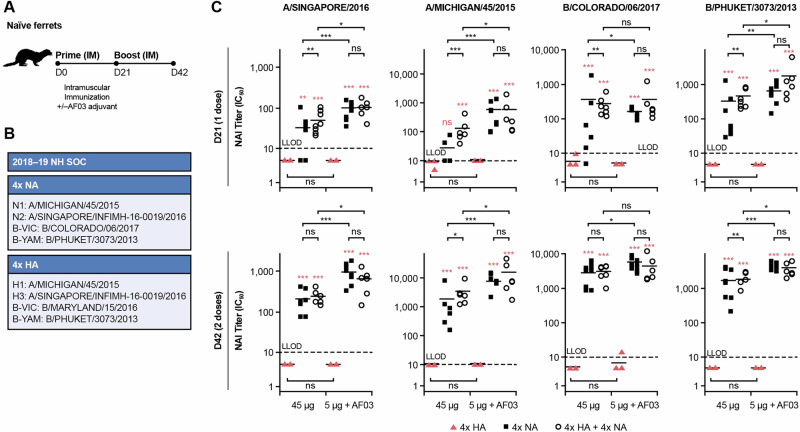
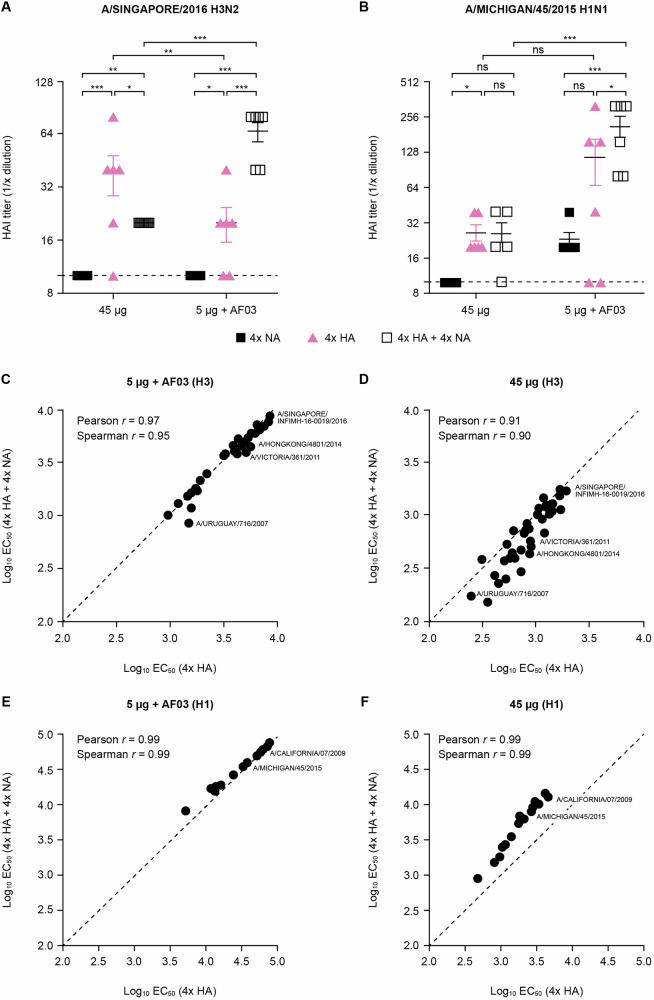
Neuraminidase (NA), the second most abundant surface glycoprotein on the influenza virus, plays a key role in viral replication and propagation. Despite growing evidence showing that NA-specific antibodies correlate with resistance to disease in humans, current licensed vaccines focus almost entirely on the hemagglutinin (HA) antigen. Here, we demonstrate that recombinant NA (rNA) protein is highly immunogenic in both naïve mice and ferrets, as well as in pre-immune ferrets, irrespective of the level of match with preexisting immunity. Ferrets vaccinated with rNA developed mild influenza disease symptoms upon challenge with human H3N2 influenza virus, and anti-NA antibody responses appeared correlated with reduction in disease severity. The addition of rNA to a quadrivalent HA-based vaccine induced robust NA-specific humoral immunity in ferrets, while retaining the ability to induce HA-specific immunity. These results demonstrate that the addition of rNA is a viable option to increase immunogenicity and potentially efficacy versus currently licensed influenza vaccines by means of boosting NA immunity.
© 2024. The Author(s).
Conflict of interest statement
Figures
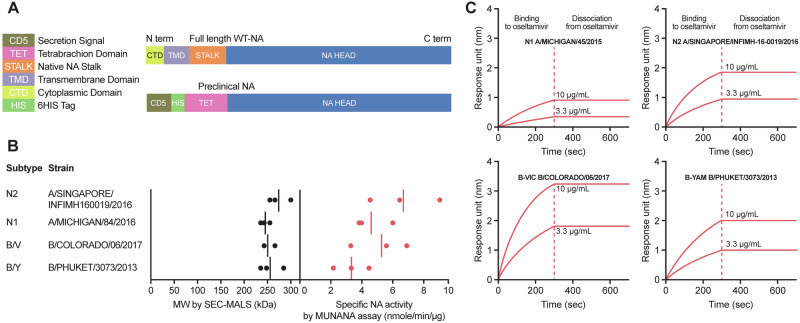
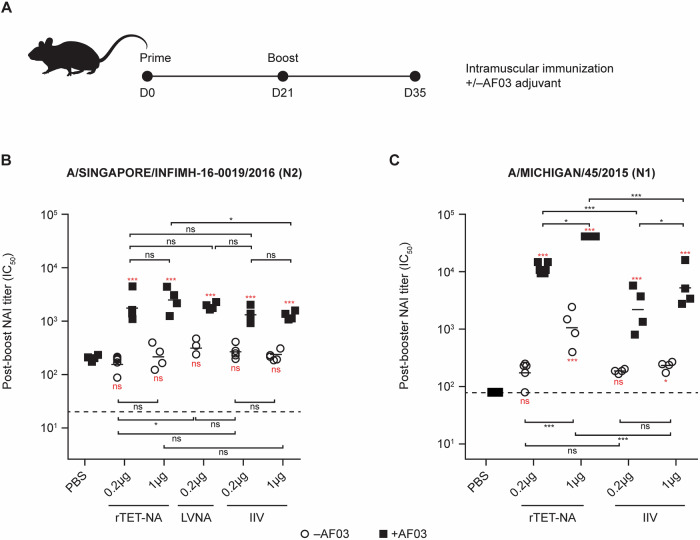
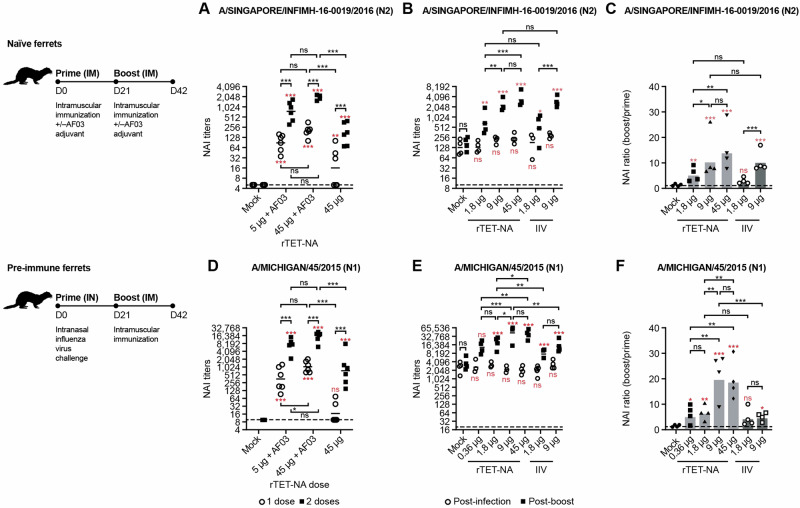
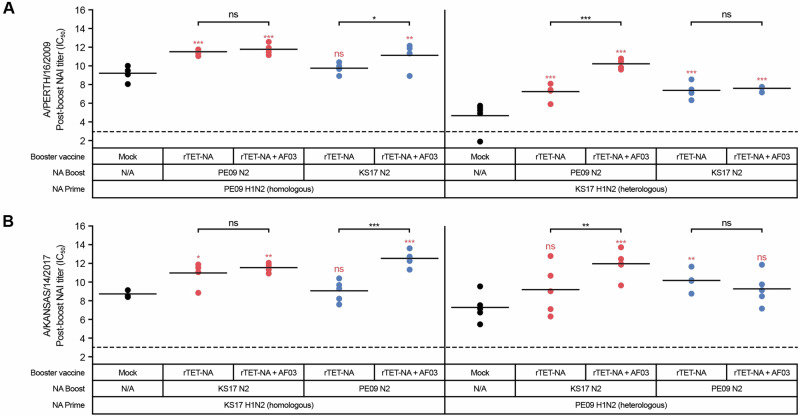
References
-
- Brueggemann, A. B. et al. Changes in the incidence of invasive disease due to Streptococcus pneumoniae, Haemophilus influenzae, and Neisseria meningitidis during the COVID-19 pandemic in 26 countries and territories in the Invasive Respiratory Infection Surveillance Initiative: a prospective analysis of surveillance data. Lancet Dig. Health3, e360–e370 (2021). - PMC - PubMed
LinkOut - more resources
Full Text Sources
