

Spatial Control over Reactions via Localized Transcription within Membraneless DNA Nanostar Droplets
- PMID: 39565729
- PMCID: PMC11701973
- DOI: 10.1021/jacs.4c07274
Spatial Control over Reactions via Localized Transcription within Membraneless DNA Nanostar Droplets
Abstract
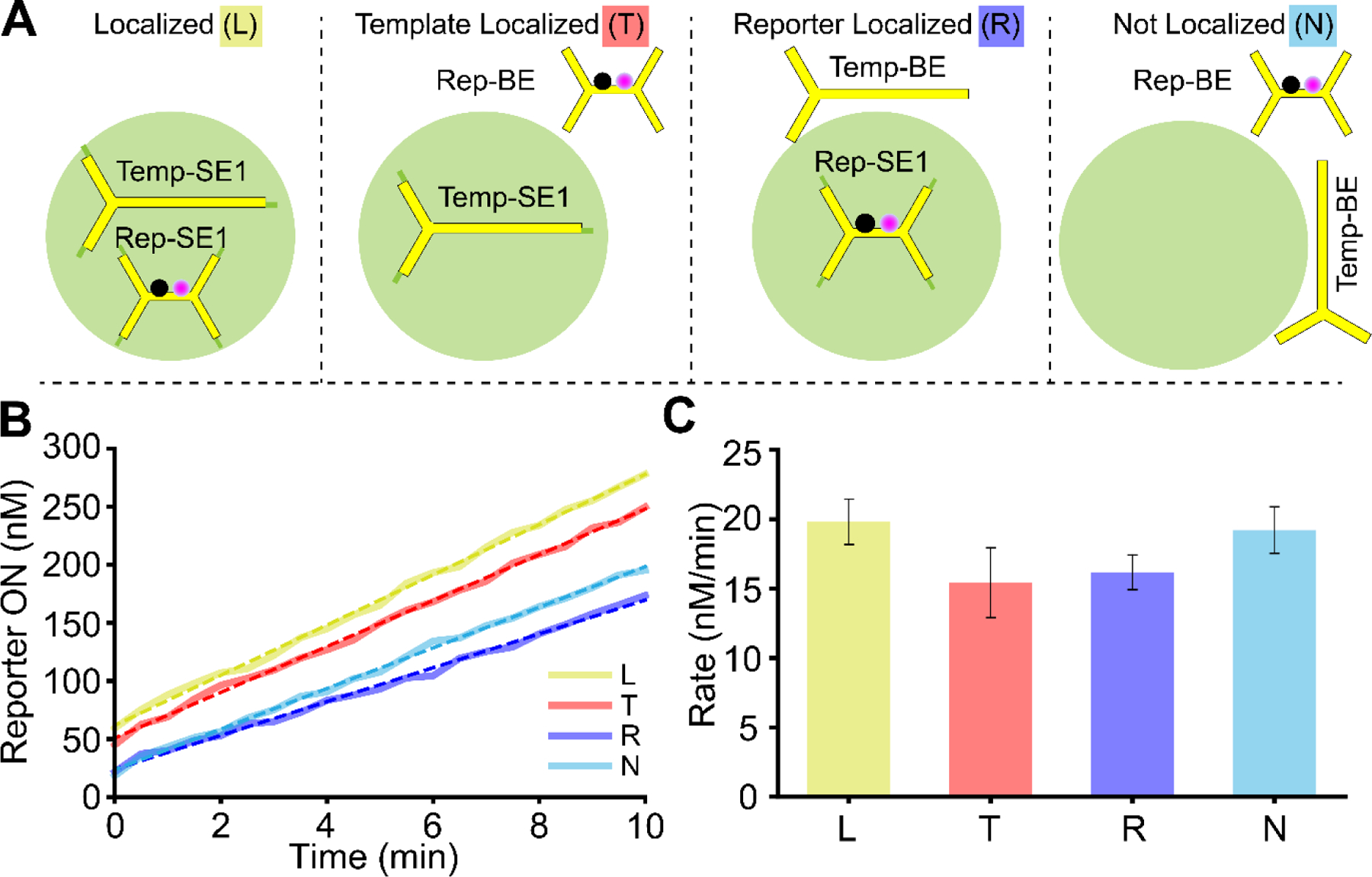
Biomolecular condensates control where and how fast many chemical reactions occur in cells by partitioning reactants and catalysts, enabling simultaneous reactions in different spatial locations of a cell. Even without a membrane or physical barrier, the partitioning of the reactants can affect the rates of downstream reaction cascades in ways that depend on reaction location. Such effects can enable systems of biomolecular condensates to spatiotemporally orchestrate chemical reaction networks in cells to facilitate complex behaviors such as ribosome assembly. Here, we develop a system for developing such control in synthetic systems. We localize different transcription templates within different phase-separated, membraneless DNA nanostar (NS) droplets─programmable, in vitro liquid-liquid phase separation systems for partitioning of substrates and localization of reactions to membraneless droplets. When RNA produced within such droplets is also degraded in the bulk, droplet-localized transcription creates RNA concentration gradients. Consistent with the formation of these gradients, toehold-mediated strand displacement reactions involving transcripts are 2-fold slower far from the site of transcription than when nearby. We then demonstrate how multiple such gradients can form and be maintained independently by simultaneous transcription reactions occurring in tandem, each localized to different NS droplet types. Our results provide a means for constructing reaction systems in which different reactions are spatially localized and controlled without the need for physical membranes. This system also provides a means for generally studying how localized reactions and the exchange of reaction products might occur between protocells.
Conflict of interest statement
Competing Interests Statement
The authors declare no competing interests.
Figures





References
-
- Tatomer DC; Terzo E; Curry KP; Salzler H; Sabath I; Zapotoczny G; McKay DJ; Dominski Z; Marzluff WF; Duronio RJ Concentrating Pre-mRNA Processing Factors in the Histone Locus Body Facilitates Efficient Histone mRNA Biogenesis. J. Cell Biol 2016, 213 (5), 557–570. 10.1083/jcb.201504043. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
