

Rotary mechanism of the prokaryotic Vo motor driven by proton motive force
- PMID: 39567487
- PMCID: PMC11579504
- DOI: 10.1038/s41467-024-53504-x
Rotary mechanism of the prokaryotic Vo motor driven by proton motive force
Abstract
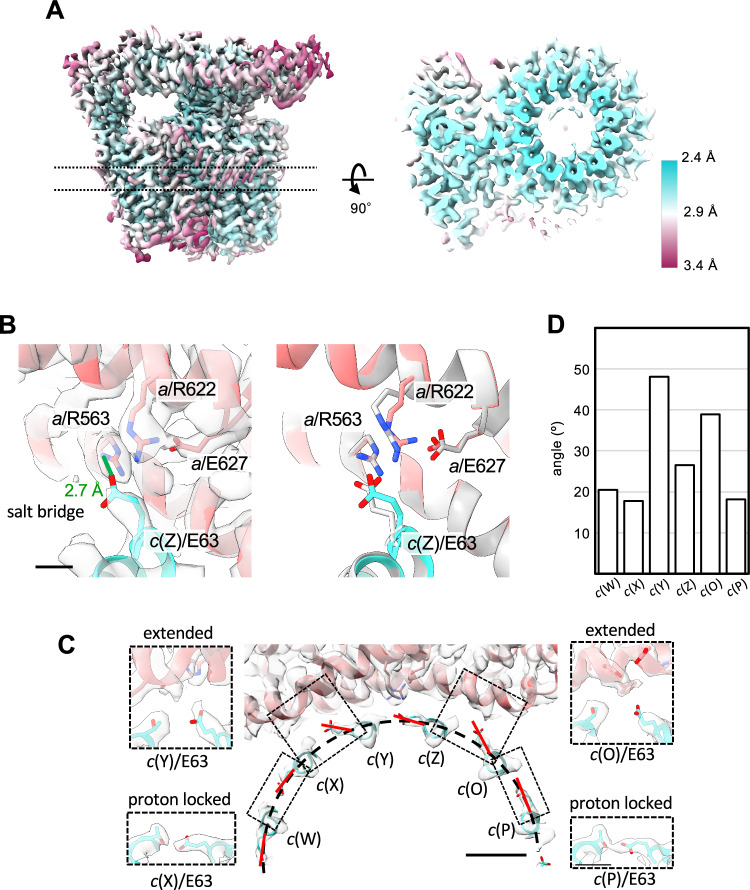
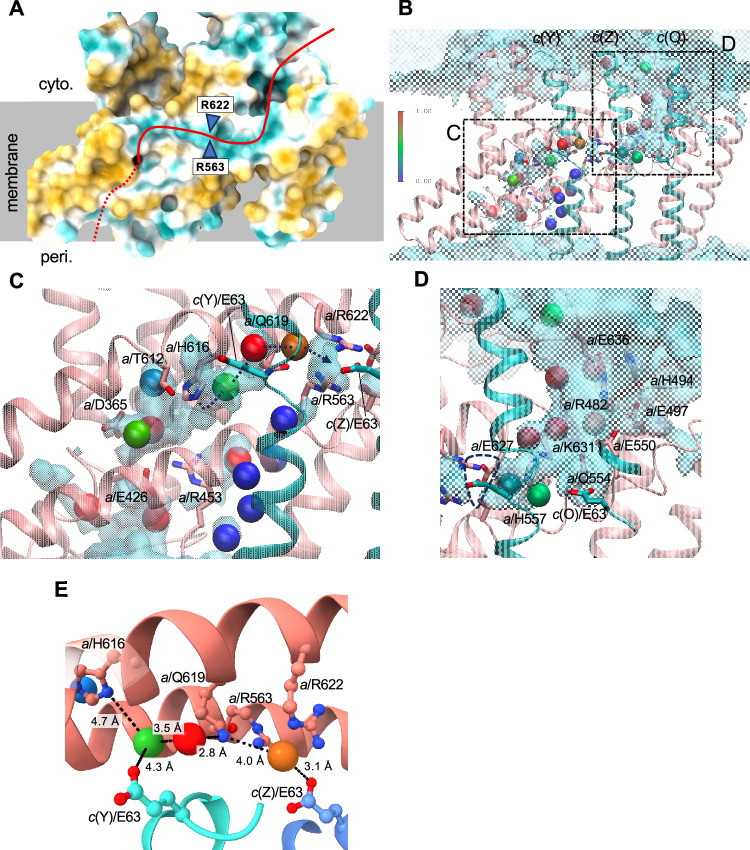
ATP synthases play a crucial role in energy production by utilizing the proton motive force (pmf) across the membrane to rotate their membrane-embedded rotor c-ring, and thus driving ATP synthesis in the hydrophilic catalytic hexamer. However, the mechanism of how pmf converts into c-ring rotation remains unclear. This study presents a 2.8 Å cryo-EM structure of the Vo domain of V/A-ATPase from Thermus thermophilus, revealing precise orientations of glutamate (Glu) residues in the c12-ring. Three Glu residues face a water channel, with one forming a salt bridge with the Arginine in the stator (a/Arg). Molecular dynamics (MD) simulations show that protonation of specific Glu residues triggers unidirectional Brownian motion of the c12-ring towards ATP synthesis. When the key Glu remains unprotonated, the salt bridge persists, blocking rotation. These findings suggest that asymmetry in the protonation of c/Glu residues biases c12-ring movement, facilitating rotation and ATP synthesis.
© 2024. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures




Similar articles
-
Cryo EM structure of intact rotary H+-ATPase/synthase from Thermus thermophilus.Nat Commun. 2018 Jan 8;9(1):89. doi: 10.1038/s41467-017-02553-6. Nat Commun. 2018. PMID: 29311594 Free PMC article.
-
Dodecamer rotor ring defines H+/ATP ratio for ATP synthesis of prokaryotic V-ATPase from Thermus thermophilus.Proc Natl Acad Sci U S A. 2007 Dec 18;104(51):20256-61. doi: 10.1073/pnas.0706914105. Epub 2007 Dec 11. Proc Natl Acad Sci U S A. 2007. PMID: 18077374 Free PMC article.
-
Atomic model for the membrane-embedded VO motor of a eukaryotic V-ATPase.Nature. 2016 Nov 3;539(7627):118-122. doi: 10.1038/nature19828. Epub 2016 Oct 24. Nature. 2016. PMID: 27776355 Free PMC article.
-
Cryo-EM studies of the structure and dynamics of vacuolar-type ATPases.Sci Adv. 2016 Jul 22;2(7):e1600725. doi: 10.1126/sciadv.1600725. eCollection 2016 Jul. Sci Adv. 2016. PMID: 27532044 Free PMC article. Review.
-
Rotation, structure, and classification of prokaryotic V-ATPase.J Bioenerg Biomembr. 2005 Dec;37(6):405-10. doi: 10.1007/s10863-005-9480-1. J Bioenerg Biomembr. 2005. PMID: 16691473 Review.
Cited by
-
Bismuth drug eradicates multi-drug resistant Burkholderia cepacia complex via aerobic respiration.Chem Sci. 2025 May 9;16(27):12372-12384. doi: 10.1039/d5sc02049b. eCollection 2025 Jul 10. Chem Sci. 2025. PMID: 40438165 Free PMC article.
-
Rotation-Direction-Dependent Mechanism of the Inhibitor Protein IF1 for Mitochondrial ATP Synthase from Atomistic Simulations.JACS Au. 2025 May 27;5(6):2654-2665. doi: 10.1021/jacsau.5c00261. eCollection 2025 Jun 23. JACS Au. 2025. PMID: 40575285 Free PMC article.
References
-
- Boyer, P. D. The binding change mechanism for ATP synthase–some probabilities and possibilities. Biochim. Biophys. Acta1140, 215–250 (1993). - PubMed
-
- Mitchell, P. Chemiosmotic coupling in oxidative and photosynthetic phosphorylation. Biol. Rev. Camb. Philos. Soc.41, 445–502 (1966). - PubMed
-
- Senior, A. E. ATP synthesis by oxidative phosphorylation. Physiol. Rev.68, 177–231 (1988). - PubMed
-
- Yoshida, M., Muneyuki, E. & Hisabori, T. ATP synthase - a marvellous rotary engine of the cell. Nat. Rev. Mol. Cell Biol.2, 669–677 (2001). - PubMed
-
- Kuhlbrandt, W. Structure and mechanisms of F-type ATP synthases. Annu. Rev. Biochem.88, 515–549 (2019). - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
