

A spatial human thymus cell atlas mapped to a continuous tissue axis
- PMID: 39567784
- PMCID: PMC11578893
- DOI: 10.1038/s41586-024-07944-6
A spatial human thymus cell atlas mapped to a continuous tissue axis
Abstract
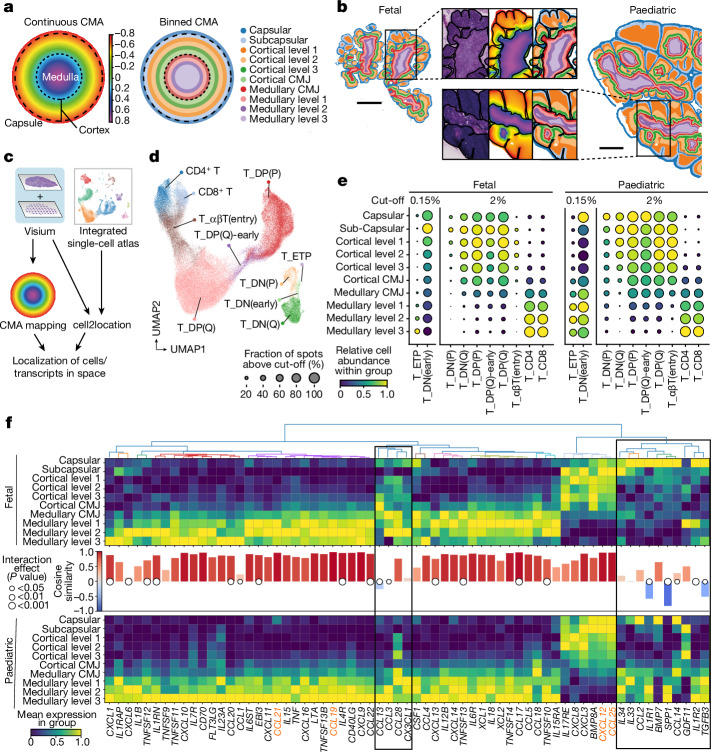
T cells develop from circulating precursor cells, which enter the thymus and migrate through specialized subcompartments that support their maturation and selection1. In humans, this process starts in early fetal development and is highly active until thymic involution in adolescence. To map the microanatomical underpinnings of this process in pre- and early postnatal stages, we established a quantitative morphological framework for the thymus-the Cortico-Medullary Axis-and used it to perform a spatially resolved analysis. Here, by applying this framework to a curated multimodal single-cell atlas, spatial transcriptomics and high-resolution multiplex imaging data, we demonstrate establishment of the lobular cytokine network, canonical thymocyte trajectories and thymic epithelial cell distributions by the beginning of the the second trimester of fetal development. We pinpoint tissue niches of thymic epithelial cell progenitors and distinct subtypes associated with Hassall's corpuscles and identify divergence in the timing of medullary entry between CD4 and CD8 T cell lineages. These findings provide a basis for a detailed understanding of T lymphocyte development and are complemented with a holistic toolkit for cross-platform imaging data analysis, annotation and OrganAxis construction (TissueTag), which can be applied to any tissue.
© 2024. The Author(s).
Conflict of interest statement
Competing interests: J.C.M. has been an employee of Genentech since September 2022. S.A.T. is a scientific advisory board member of ForeSite Labs, OMass Therapeutics, Qiagen, a co-founder and equity holder of TransitionBio and EnsoCell Therapeutics, a non-executive director of 10x Genomics and a part-time employee of GlaxoSmithKline. The other authors declare no competing interests.
Figures













Update of
-
A spatial human thymus cell atlas mapped to a continuous tissue axis.bioRxiv [Preprint]. 2023 Oct 27:2023.10.25.562925. doi: 10.1101/2023.10.25.562925. bioRxiv. 2023. Update in: Nature. 2024 Nov;635(8039):708-718. doi: 10.1038/s41586-024-07944-6. PMID: 37986877 Free PMC article. Updated. Preprint.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
