

An integrated transcriptomic cell atlas of human neural organoids
- PMID: 39567792
- PMCID: PMC11578878
- DOI: 10.1038/s41586-024-08172-8
An integrated transcriptomic cell atlas of human neural organoids
Erratum in
-
Publisher Correction: An integrated transcriptomic cell atlas of human neural organoids.Nature. 2025 Jan;637(8045):E21. doi: 10.1038/s41586-024-08482-x. Nature. 2025. PMID: 39663459 Free PMC article. No abstract available.
Abstract
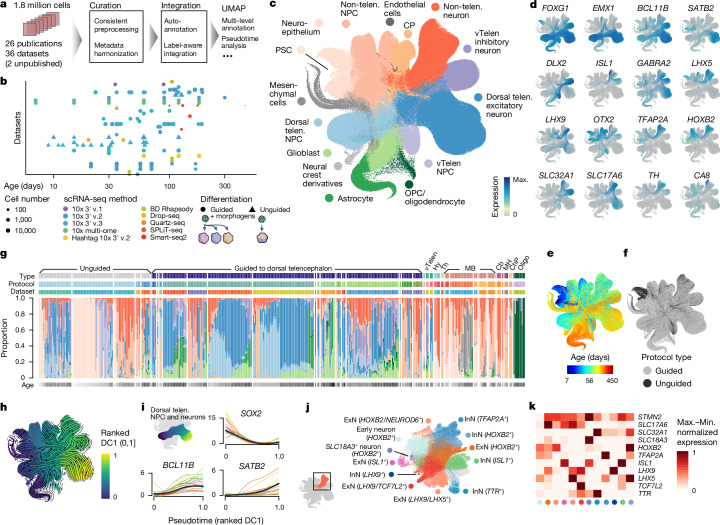
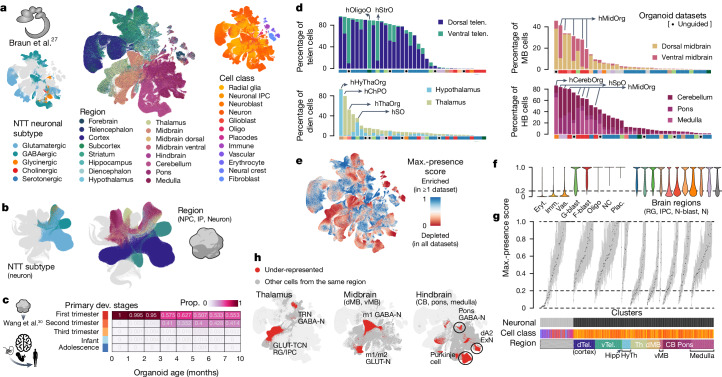
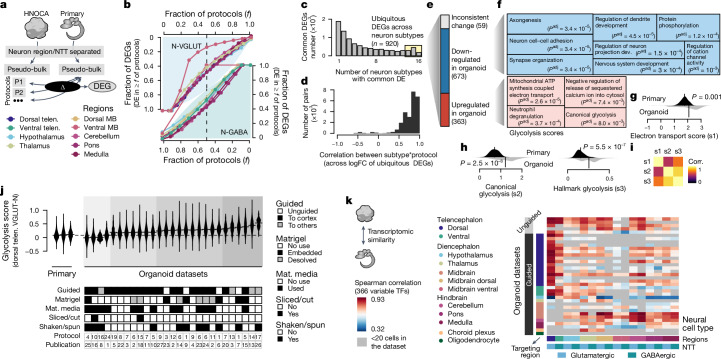
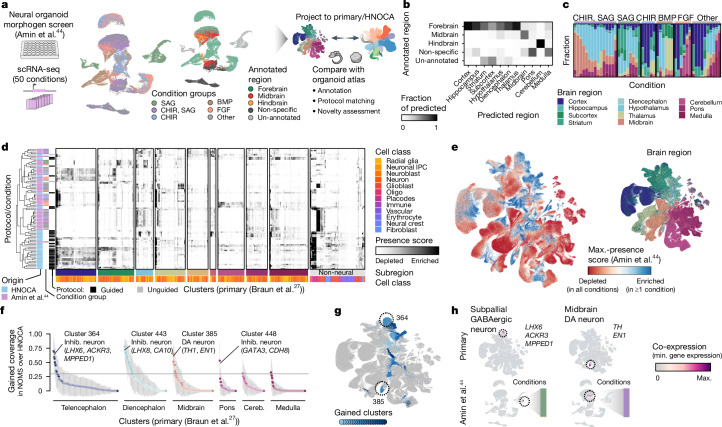
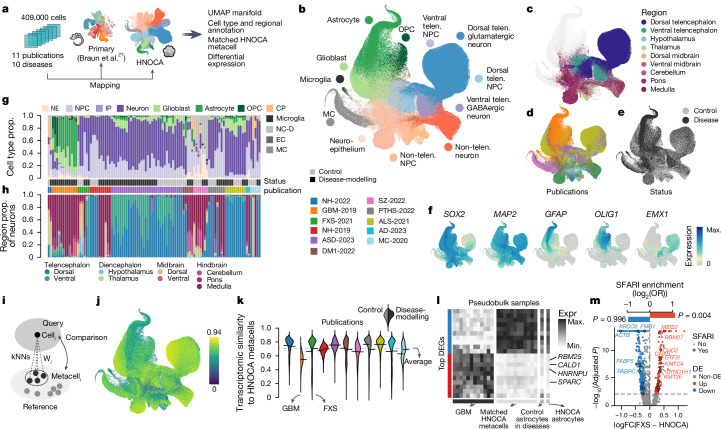
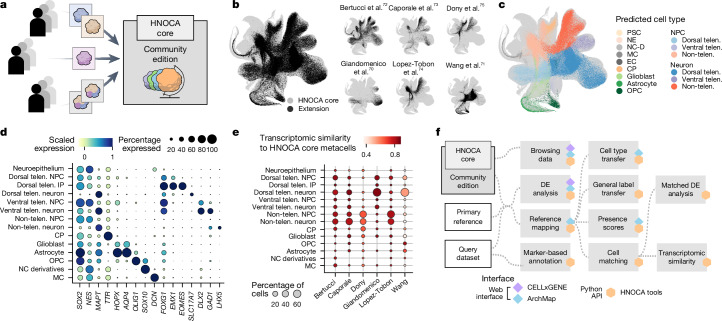
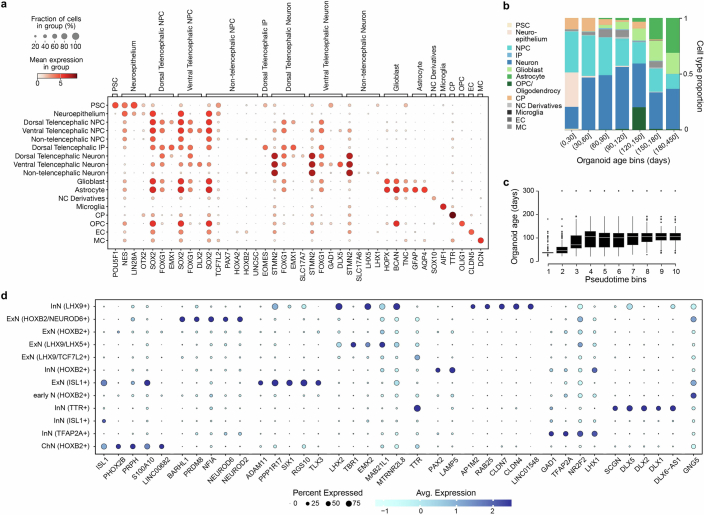
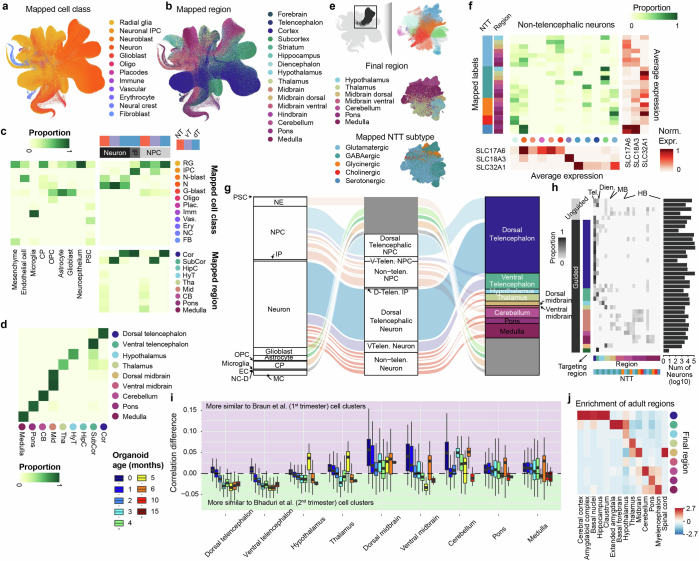
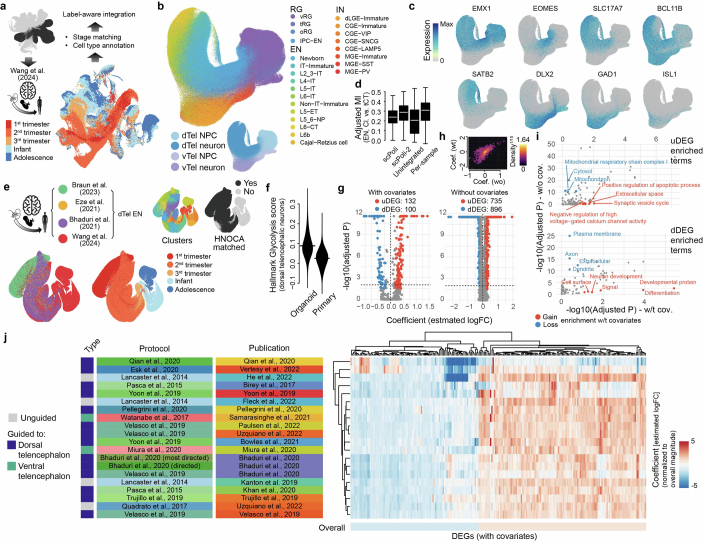
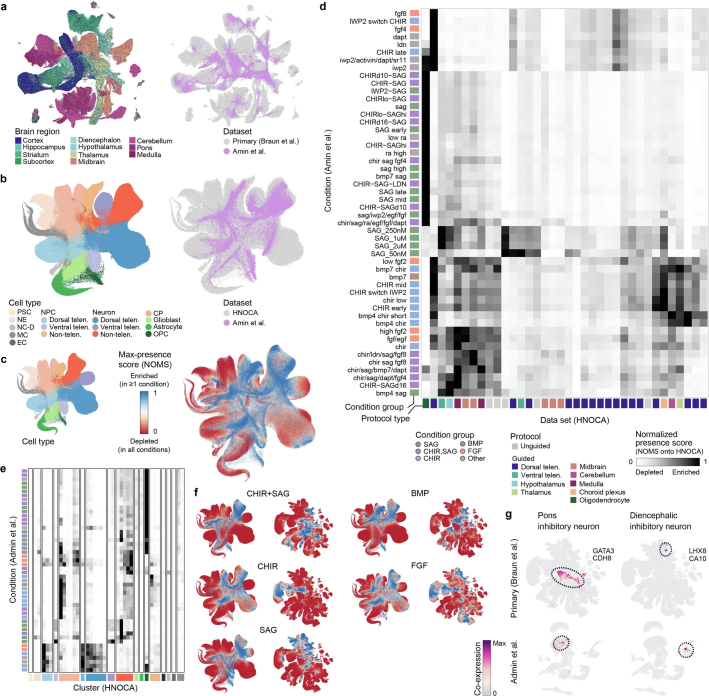
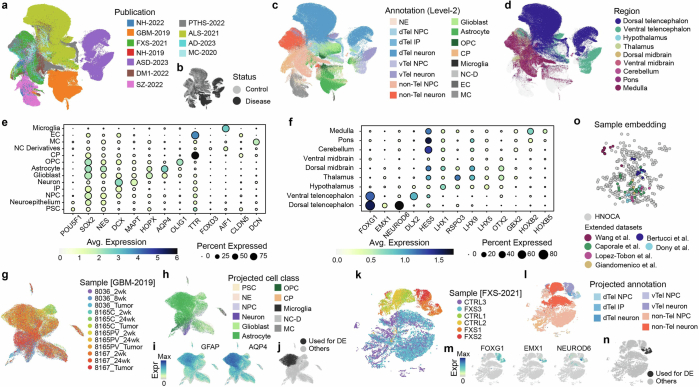
Human neural organoids, generated from pluripotent stem cells in vitro, are useful tools to study human brain development, evolution and disease. However, it is unclear which parts of the human brain are covered by existing protocols, and it has been difficult to quantitatively assess organoid variation and fidelity. Here we integrate 36 single-cell transcriptomic datasets spanning 26 protocols into one integrated human neural organoid cell atlas totalling more than 1.7 million cells1-26. Mapping to developing human brain references27-30 shows primary cell types and states that have been generated in vitro, and estimates transcriptomic similarity between primary and organoid counterparts across protocols. We provide a programmatic interface to browse the atlas and query new datasets, and showcase the power of the atlas to annotate organoid cell types and evaluate new organoid protocols. Finally, we show that the atlas can be used as a diverse control cohort to annotate and compare organoid models of neural disease, identifying genes and pathways that may underlie pathological mechanisms with the neural models. The human neural organoid cell atlas will be useful to assess organoid fidelity, characterize perturbed and diseased states and facilitate protocol development.
© 2024. The Author(s).
Conflict of interest statement
Competing interests: F.J.T. consults for Immunai Inc., Singularity Bio B.V., CytoReason Ltd, Cellarity, and has ownership interest in Dermagnostix GmbH and Cellarity. The other authors declare no competing interests.
Figures

References
-
- Kanton, S. et al. Organoid single-cell genomic atlas uncovers human-specific features of brain development. Nature574, 418–422 (2019). - PubMed
MeSH terms
LinkOut - more resources
Full Text Sources
