

Plasmodium berghei liver stage parasites exploit host GABARAP proteins for TFEB activation
- PMID: 39572689
- PMCID: PMC11582615
- DOI: 10.1038/s42003-024-07242-x
Plasmodium berghei liver stage parasites exploit host GABARAP proteins for TFEB activation
Abstract
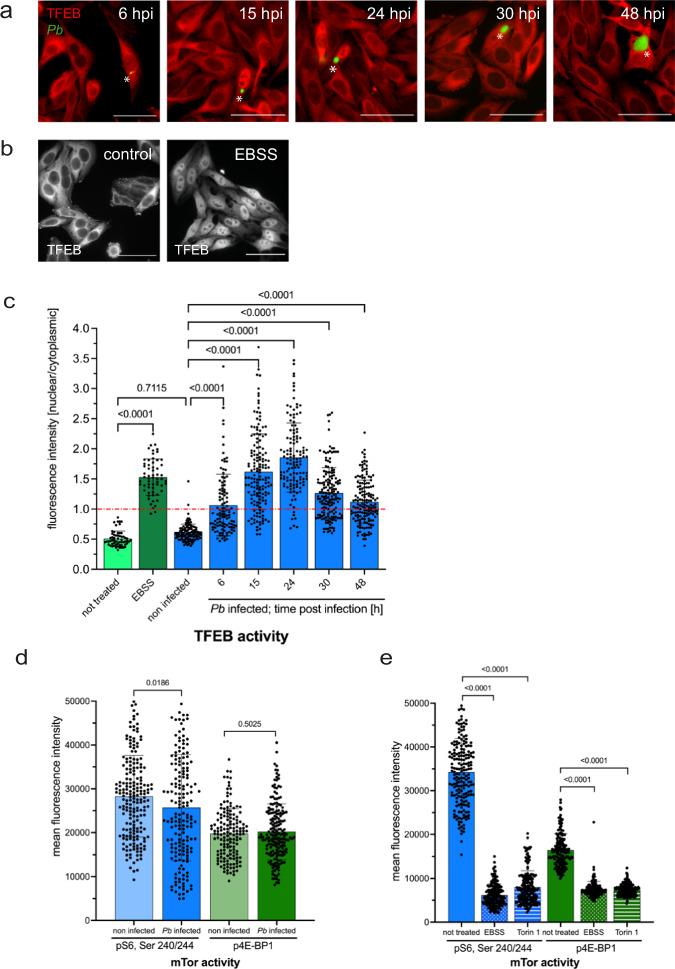
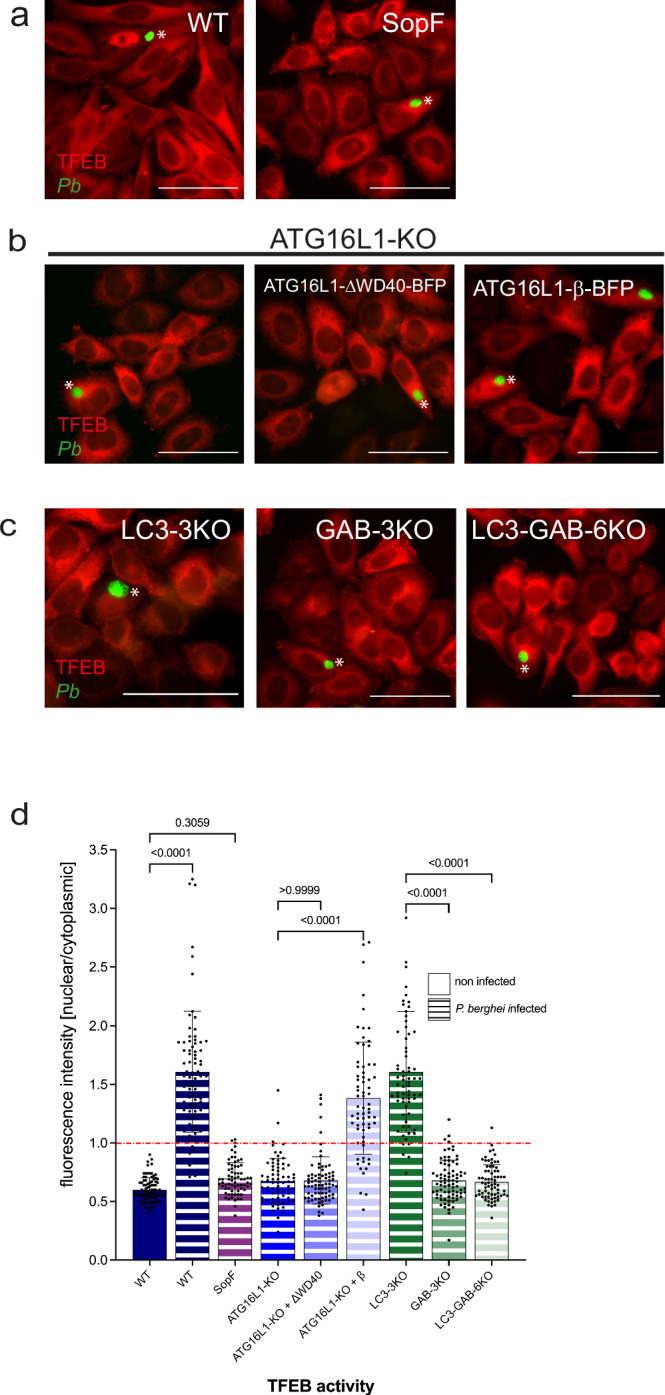
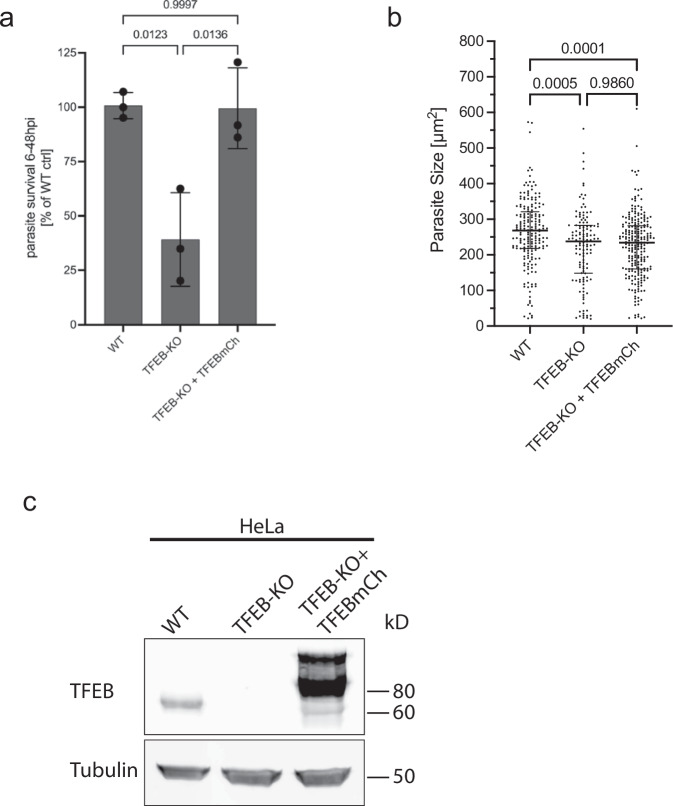
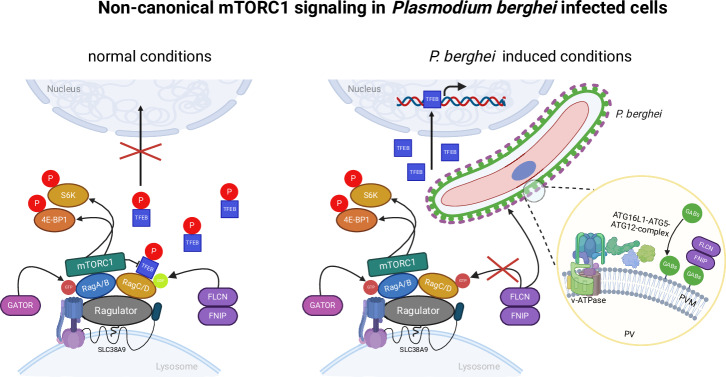
Plasmodium, the causative agent of malaria, infects hepatocytes prior to establishing a symptomatic blood stage infection. During this liver stage development, parasites reside in a parasitophorous vacuole (PV), whose membrane acts as the critical interface between the parasite and the host cell. It is well-established that host cell autophagy-related processes significantly impact the development of Plasmodium liver stages. Expression of genes related to autophagy and lysosomal biogenesis is orchestrated by transcription factor EB (TFEB). In this study, we explored the activation of host cell TFEB in Plasmodium berghei-infected cells during the liver stage of the parasite. Our results unveiled a critical role of proteins belonging to the Gamma-aminobutyric acid receptor-associated protein subfamily (GABARAP) of ATG8 proteins (GABARAP/L1/L2 and LC3A/B/C) in recruiting the TFEB-blocking FLCN-FNIP (Folliculin-Folliculin-interacting protein) complex to the PVM. Remarkably, the sequestration of FLCN-FNIP resulted in a robust activation of TFEB, reliant on conjugation of ATG8 proteins to single membranes (CASM) and GABARAP proteins. Our findings provide novel mechanistic insights into host cell signaling occurring at the PVM, shedding light on the complex interplay between Plasmodium parasites and the host cell during the liver stage of infection.
© 2024. The Author(s).
Conflict of interest statement
Competing interests: The authors declare the following competing interests: L.O.M. and T.L. are employees and shareholders of Casma Therapeutics. A.B. is a co-founder and shareholder of CASMA Therapeutics, Inc, and Advisory board member of Avilar Therapeutics and Amplify Therapeutics. L.O.M. is an author on a patent related to TFEB modulation, in addition. M.L. is a co-founder and member of the scientific advisory board of Automera. Other authors declare no competing interests.
Figures


References
-
- Frischknecht, F. et al. Imaging movement of malaria parasites during transmission by Anopheles mosquitoes. Cell. Microbiol.6, 687–694 (2004). - PubMed
-
- Amino, R. et al. Quantitative imaging of Plasmodium transmission from mosquito to mammal. Nat. Med.12, 220–224 (2006). - PubMed
-
- Pradel, G. & Frevert, U. Malaria sporozoites actively enter and pass through rat Kupffer cells prior to hepatocyte invasion. Hepatology33, 1154–1165 (2001). - PubMed
-
- Sturm, A. et al. Manipulation of host hepatocytes by the malaria parasite for delivery into liver sinusoids. Science313, 1287–1290 (2006). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
