

This is a preprint.
Suppression of Huntington's Disease Somatic Instability by Transcriptional Repression and Direct CAG Repeat Binding
- PMID: 39574582
- PMCID: PMC11580907
- DOI: 10.1101/2024.11.04.619693
Suppression of Huntington's Disease Somatic Instability by Transcriptional Repression and Direct CAG Repeat Binding
Update in
-
Suppression of Huntington's Disease Somatic Instability by Transcriptional Repression and Direct CAG Repeat Binding.Nat Commun. 2025 Nov 14;16(1):10009. doi: 10.1038/s41467-025-64936-4. Nat Commun. 2025. PMID: 41238535 Free PMC article.
Abstract
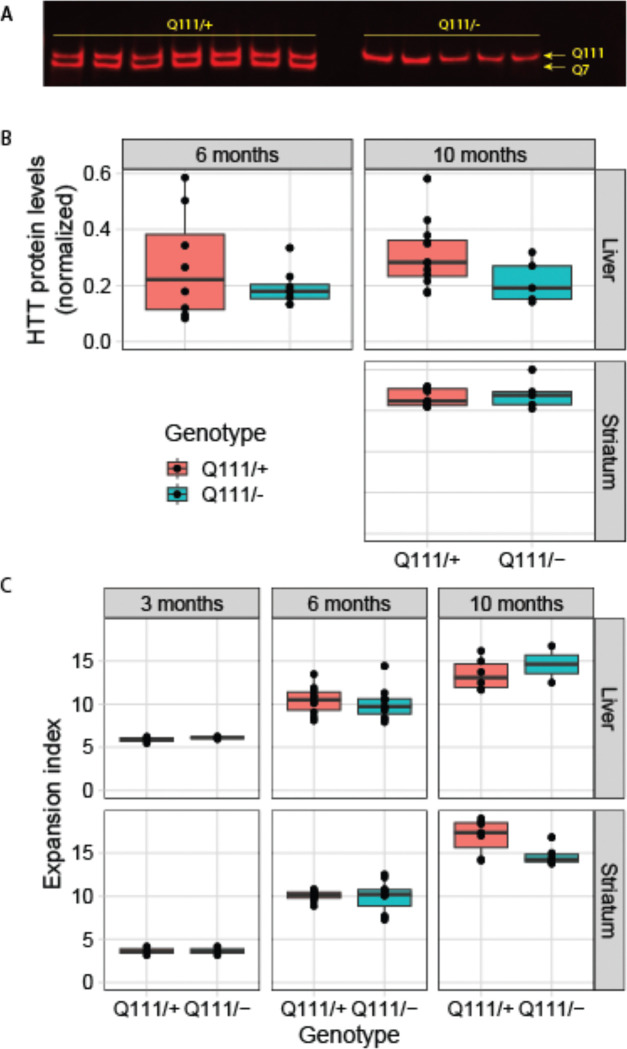
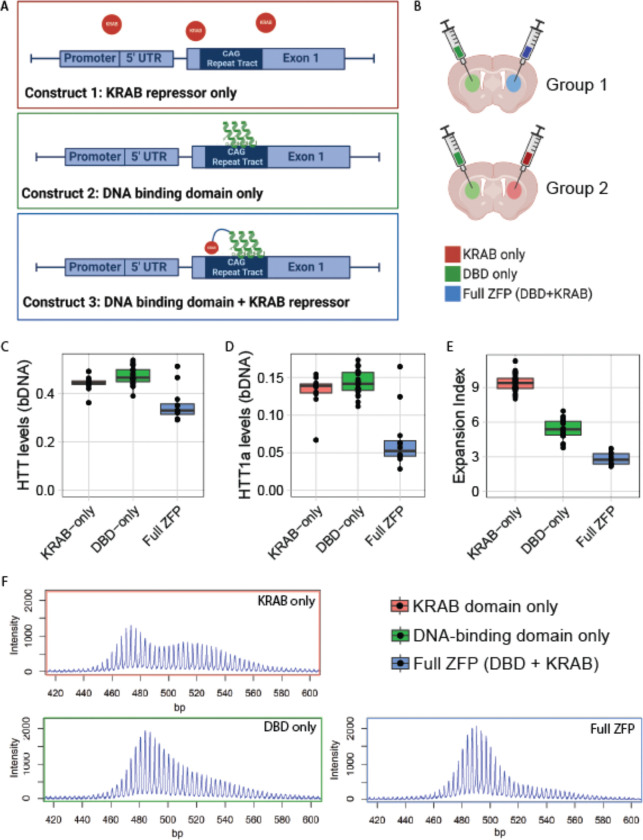
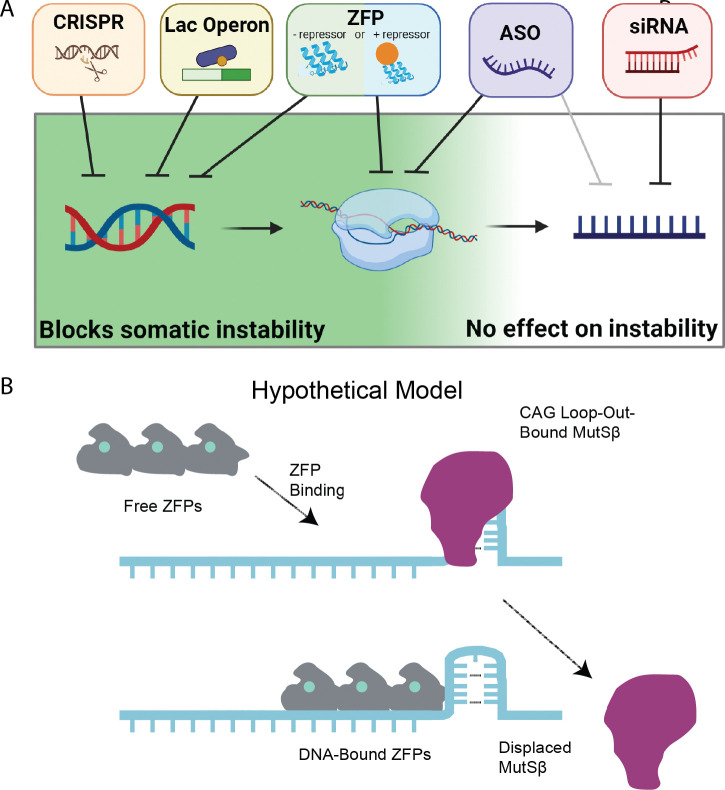
Huntington's disease (HD) arises from a CAG expansion in the huntingtin (HTT) gene beyond a critical threshold. A major thrust of current HD therapeutic development is lowering levels of mutant HTT mRNA (mHTT) and protein (mHTT) with the aim of reducing the toxicity of these product(s). Human genetic data also support a key role for somatic instability (SI) in HTT's CAG repeat - whereby it lengthens with age in specific somatic cell types - as a key driver of age of motor dysfunction onset. Thus, an attractive HD therapy would address both mHTT toxicity and SI, but to date the relationship between SI and HTT lowering remains unexplored. Here, we investigated multiple therapeutically-relevant HTT-lowering modalities to establish the relationship between HTT lowering and SI in HD knock-in mice. We find that repressing transcription of mutant Htt (mHtt) provides robust protection from SI, using diverse genetic and pharmacological approaches (antisense oligonucleotides, CRISPR-Cas9 genome editing, the Lac repressor, and virally delivered zinc finger transcriptional repressor proteins, ZFPs). However, we find that small interfering RNA (siRNA), a potent HTT-lowering treatment, lowers HTT levels without influencing SI and that SI is also normal in mice lacking 50% of total HTT levels, suggesting HTT levels, per se, do not modulate SI in trans. Remarkably, modified ZFPs that bind the mHtt locus, but lack a repressive domain, robustly protect from SI, despite not reducing HTT mRNA or protein levels. These results have important therapeutic implications in HD, as they suggest that DNA-targeted HTT-lowering treatments may have significant advantages compared to other HTT-lowering approaches, and that interaction of a DNA-binding protein and HTT's CAG repeats may provide protection from SI while sparing HTT expression.
Conflict of interest statement
Declaration of Interests A.N., A.S., J.F., T.S., A.C., I.L. are Employees of Evotec, and may have stock options. C.F.B. and H.K. Are full time employees at, and hold shares in, Ionis Pharmaceuticals. V.C.W. was a founding scientific advisory board member with a financial interest in Triplet Therapeutics Inc., her financial interests were reviewed and are managed by Massachusetts General Hospital (MGH) and Mass General Brigham (MGB) in accordance with their conflict of interest policies. V.C.W. is a scientific advisory board member of LoQus23 Therapeutics Ltd. and has provided paid consulting services to Acadia Pharmaceuticals Inc., Alnylam Inc., Biogen Inc., Passage Bio and Rgenta Therapeutics and has received research support from Pfizer Inc. J.B.C. Has provided paid consulting and/or conducted sponsored research for Wave Life Sciences, Skyhawk Therapeutics, Cajal Neuroscience, Ionis Pharmaceuticals, and Alnylam, and Gudiepoint. D.H., D.M. and T.V.are full-time employees of CHDI Foundation. A.K. discloses ownership of stocks in RXi Pharmaceuticals and Advirna, and is a founder of Atalanta Therapeutics and Comanche Biopharma. R.B. received consulting fees from Takeda.
Figures
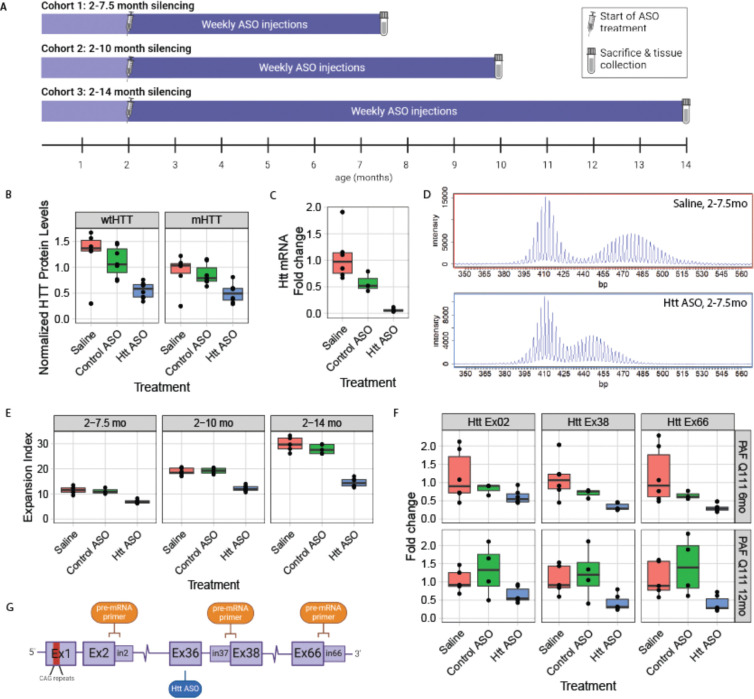
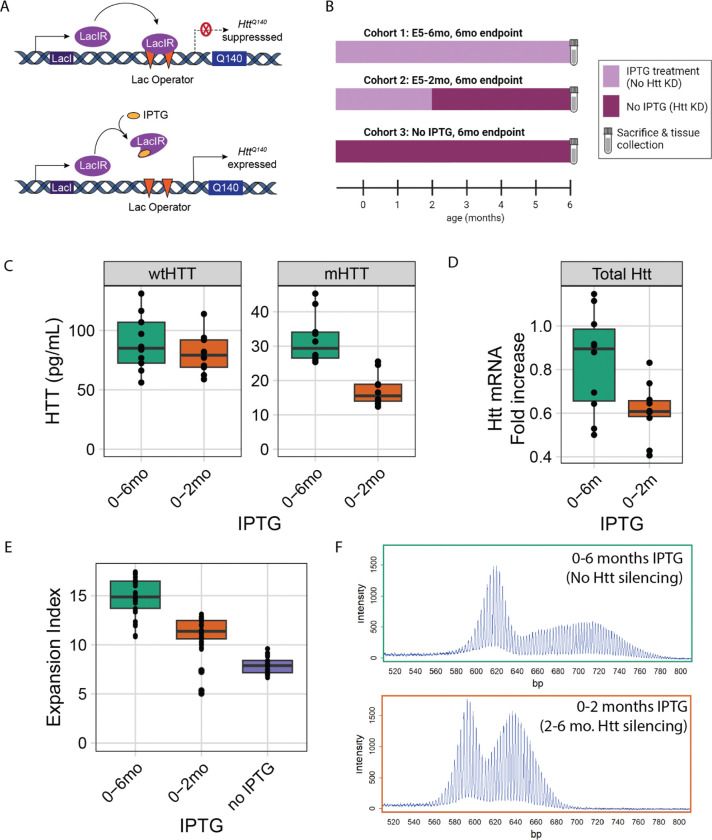
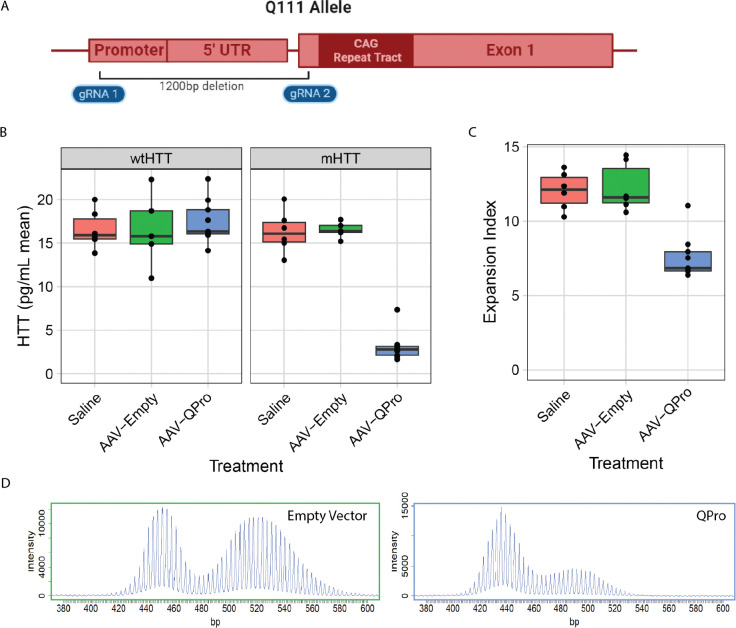
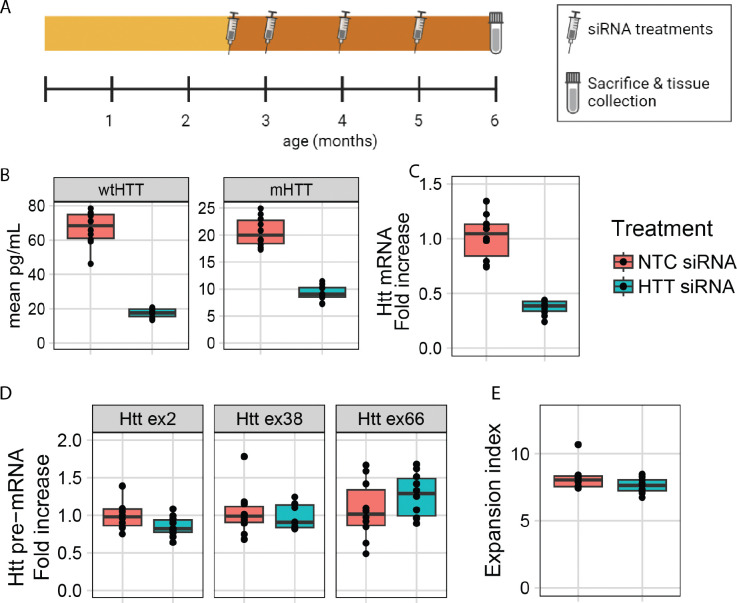
References
-
- The Huntington’s Disease Collaborative Research Group. A novel gene containing a trinucleotide repeat that is expanded and unstable on Huntington’s disease chromosomes. Cell 72, 971–83 (1993). - PubMed
-
- McColgan P. & Tabrizi S. J. Huntington’s disease: a clinical review. Eur. J. Neurol. 25, 24–34 (2018). - PubMed
-
- Andrew S. E. et al. The relationship between trinucleotide (CAG) repeat length and clinical features of Huntington’s disease. Nat. Gen. 4, 398–403 (1993). - PubMed
-
- Duyao M. et al. Trinucleotide repeat length instability and age of onset in Huntington’s disease. Nat. Gen. 4, 387–392 (1993). - PubMed
-
- Snell R. G. et al. Relationship between trinucleotide repeat expansion and phenotypic variation in Huntington’s disease. Nat. Gen. 4, 393–397 (1993). - PubMed
Publication types
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Medical