

This is a preprint.
Improved precision, sensitivity, and adaptability of Ordered Two-Template Relay cDNA library preparation for RNA sequencing
- PMID: 39574714
- PMCID: PMC11581009
- DOI: 10.1101/2024.11.09.622813
Improved precision, sensitivity, and adaptability of Ordered Two-Template Relay cDNA library preparation for RNA sequencing
Update in
-
Improved precision, sensitivity, and adaptability of ordered two-template relay cDNA library preparation for RNA sequencing.RNA. 2025 Jan 22;31(2):224-244. doi: 10.1261/rna.080318.124. RNA. 2025. PMID: 39626888 Free PMC article.
Abstract
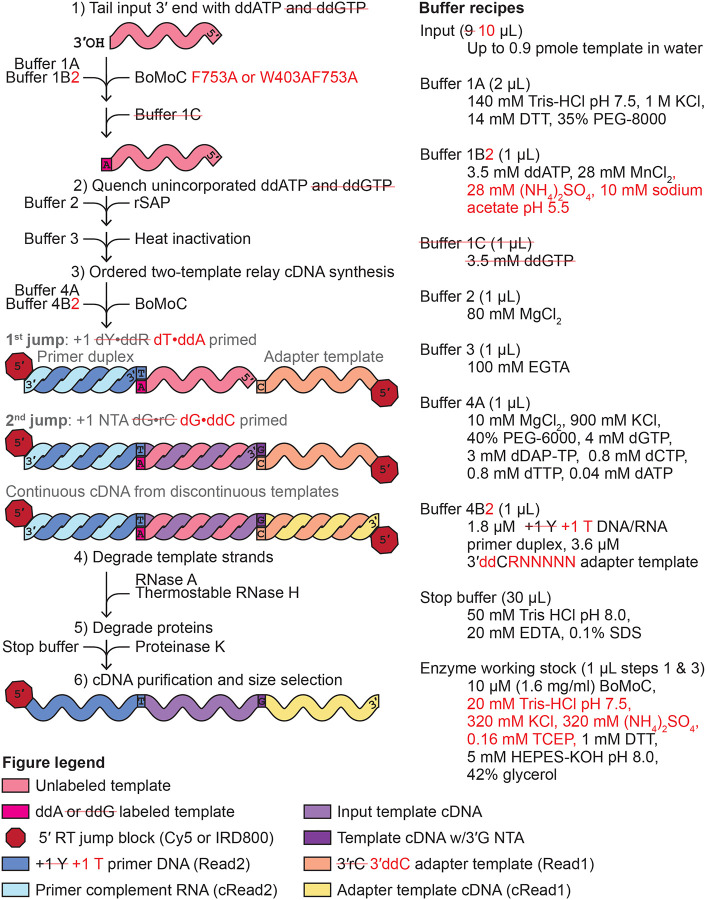
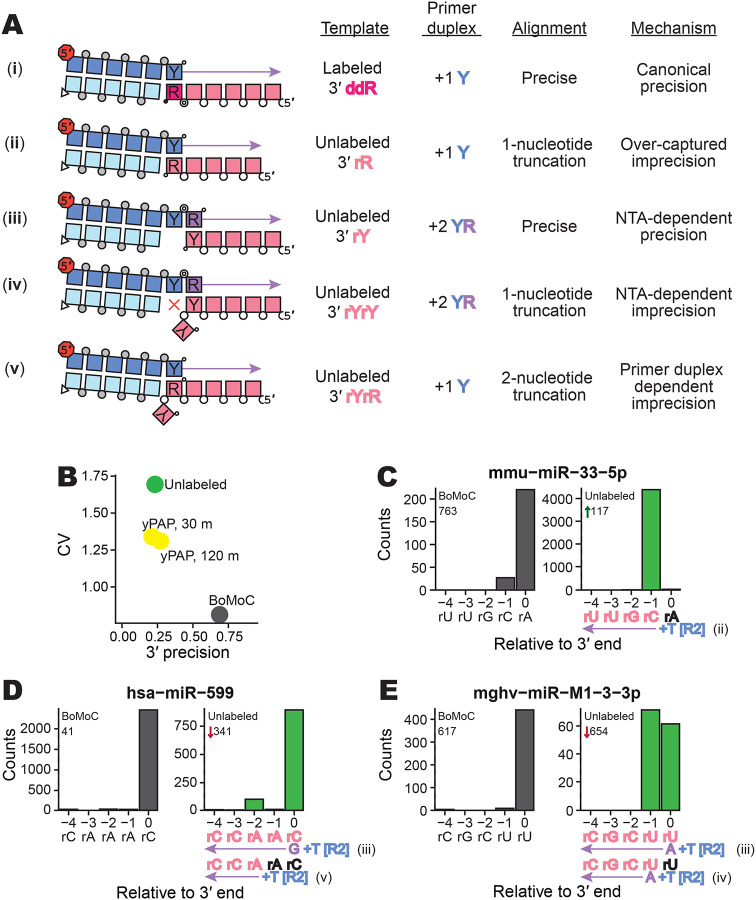
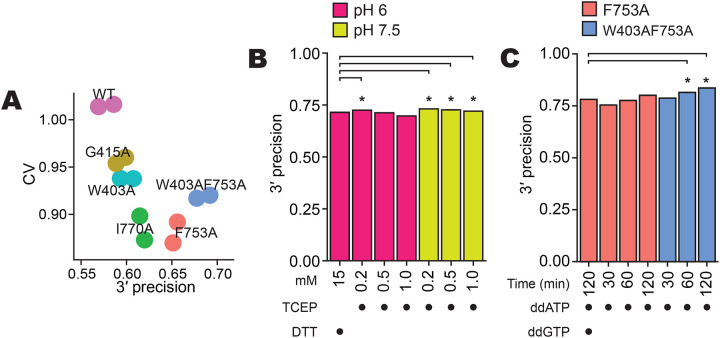
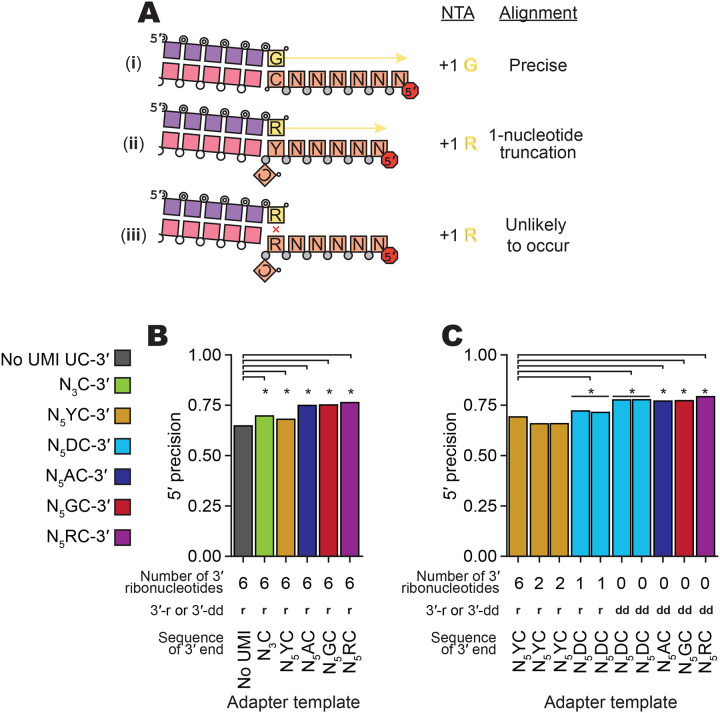
Sequencing RNAs that are biologically processed or degraded to less than ~100 nucleotides typically involves multi-step, low-yield protocols with bias and information loss inherent to ligation and/or polynucleotide tailing. We recently introduced Ordered Two-Template Relay (OTTR), a method that captures obligatorily end-to-end sequences of input molecules and, in the same reverse transcription step, also appends 5' and 3' sequencing adapters of choice. OTTR has been thoroughly benchmarked for optimal production of microRNA, tRNA and tRNA fragments, and ribosome-protected mRNA footprint libraries. Here we sought to characterize, quantify, and ameliorate any remaining bias or imprecision in the end-to-end capture of RNA sequences. We introduce new metrics for the evaluation of sequence capture and use them to optimize reaction buffers, reverse transcriptase sequence, adapter oligonucleotides, and overall workflow. Modifications of the reverse transcriptase and adapter oligonucleotides increased the 3' and 5' end-precision of sequence capture and minimized overall library bias. Improvements in recombinant expression and purification of the truncated Bombyx mori R2 reverse transcriptase used in OTTR reduced non-productive sequencing reads by minimizing bacterial nucleic acids that compete with low-input RNA molecules for cDNA synthesis, such that with miRNA input of 3 picograms (less than 1 fmol), fewer than 10% of sequencing reads are bacterial nucleic acid contaminants. We also introduce a rapid, automation-compatible OTTR protocol that enables gel-free, length-agnostic enrichment of cDNA duplexes from unwanted adapter-only side products. Overall, this work informs considerations for unbiased end-to-end capture and annotation of RNAs independent of their sequence, structure, or post-transcriptional modifications.
Keywords: OTTR; non-coding RNA; non-templated nucleotide addition; reverse transcriptase; template jumping; terminal transferase.
Conflict of interest statement
Competing Interests L.F, H.E.U., S.C.P, and K.C. are named inventors on patent applications filed by the University of California describing biochemical activities of BoMoC enzymes used for OTTR. L.F., H.E.U., and K.C. have equity in Karnateq, Inc., which licensed the technology and has produced kits for OTTR cDNA library preparation.
Figures
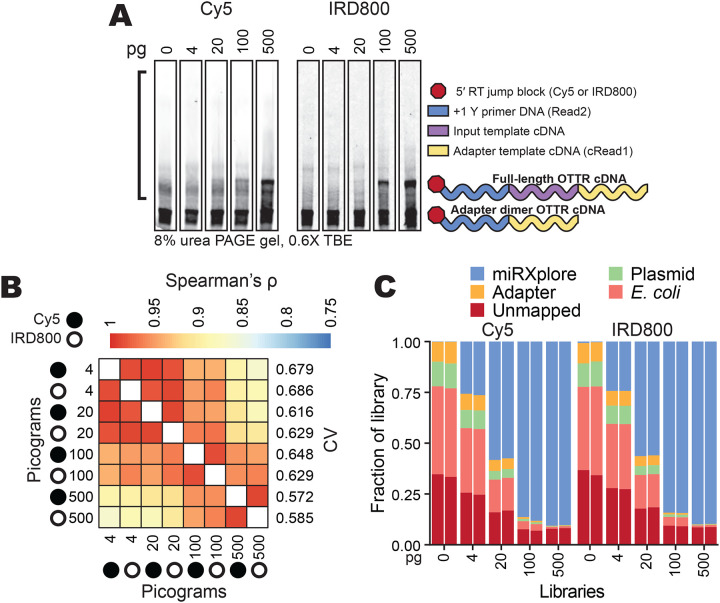
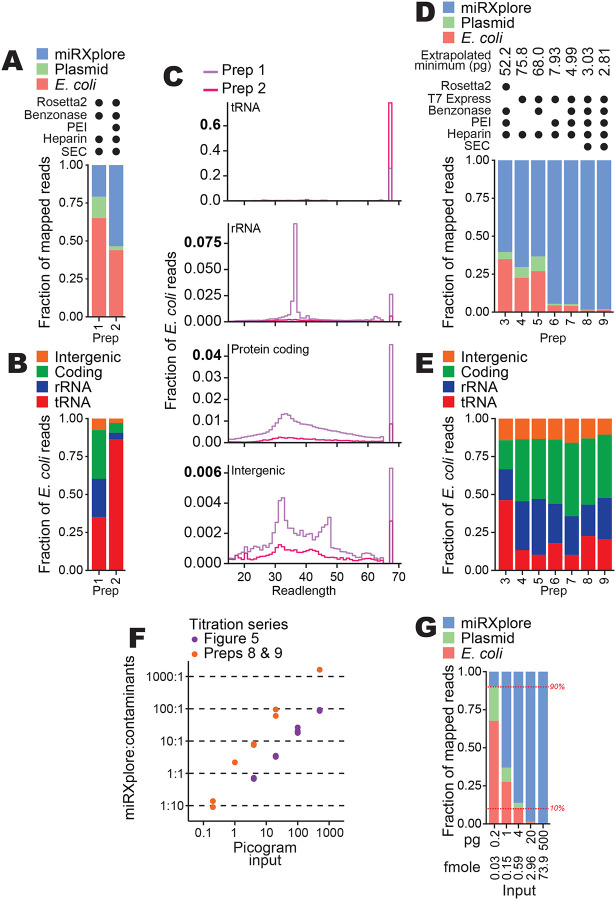
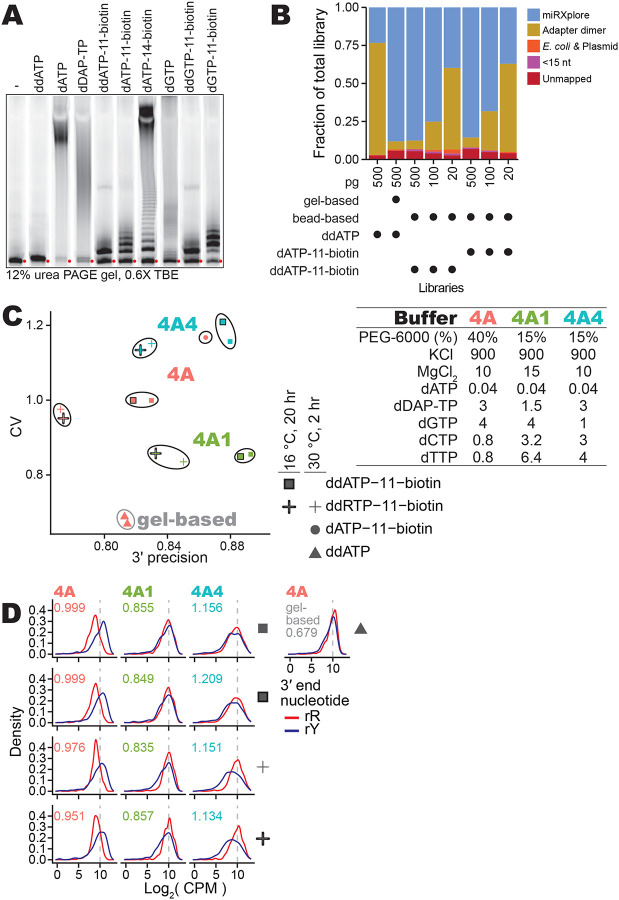
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources