

This is a preprint.
A sensitive period for the development of episodic-like memory in mice
- PMID: 39574753
- PMCID: PMC11580884
- DOI: 10.1101/2024.11.06.622296
A sensitive period for the development of episodic-like memory in mice
Update in
-
A sensitive period for the development of episodic-like memory in mice.Curr Biol. 2025 May 5;35(9):2032-2048.e3. doi: 10.1016/j.cub.2025.03.032. Epub 2025 Apr 10. Curr Biol. 2025. PMID: 40215964
Abstract
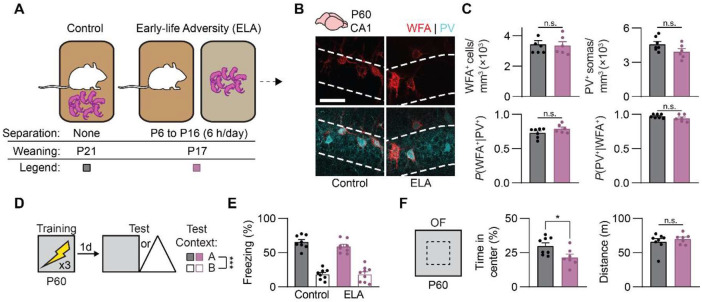
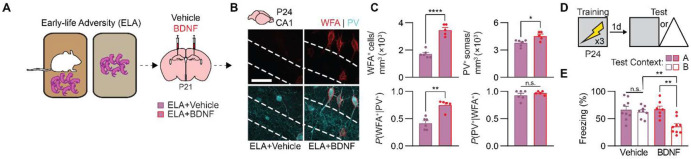
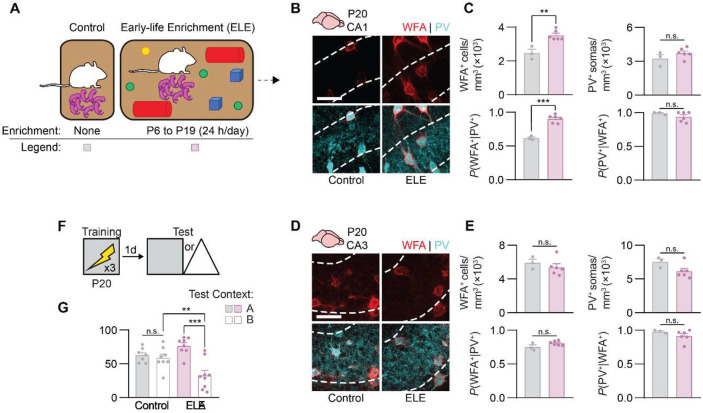
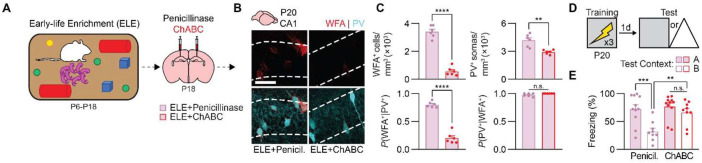
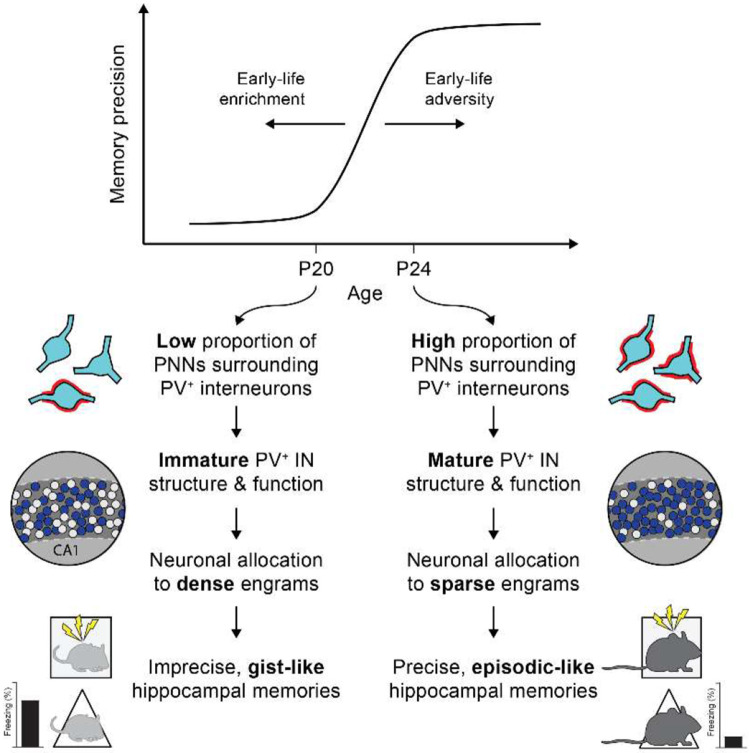
Episodic-like memory is a later-developing cognitive function supported by the hippocampus. In mice, the formation of extracellular perineuronal nets in subfield CA1 of the dorsal hippocampus controls the emergence of episodic-like memory during the fourth postnatal week (Ramsaran et al., 2023). Whether the timing of episodic-like memory onset is hard-wired, or flexibly set by early-life experiences during a critical or sensitive period for hippocampal maturation, is unknown. Here, we show that the trajectories for episodic-like memory development vary for mice given different sets of experiences spanning the second and third postnatal weeks. Specifically, episodic-like memory precision developed later in mice that experienced early-life adversity, while it developed earlier in mice that experienced early-life enrichment. Moreover, we demonstrate that early-life experiences set the timing of episodic-like memory development by modulating the pace of perineuronal net formation in dorsal CA1. These results indicate that the hippocampus undergoes a sensitive period during which early-life experiences determine the timing for episodic-like memory development.
Figures



References
-
- Akers K. G., Martinez-Canabal A., Restivo L., Yiu A. P., De Cristofaro A., Hsiang H.-L. (Liz), Wheeler A. L. Guskjolen A. Niibori Y. Shoji H. Ohira K. Richards B. A. Miyakawa T. Josselyn S. A. & Frankland P. W. (2014). Hippocampal Neurogenesis Regulates Forgetting During Adulthood and Infancy. Science, 344(6184), 598–602. 10.1126/science.1248903 - DOI - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous