

This is a preprint.
The circadian clock regulates receptor-mediated immune responses to an herbivore-associated molecular pattern
- PMID: 39574761
- PMCID: PMC11581021
- DOI: 10.1101/2024.11.06.622352
The circadian clock regulates receptor-mediated immune responses to an herbivore-associated molecular pattern
Update in
-
The Circadian Clock Regulates Receptor-Mediated Immune Responses to a Herbivore-Associated Molecular Pattern.Plant Cell Environ. 2025 Oct 23. doi: 10.1111/pce.70223. Online ahead of print. Plant Cell Environ. 2025. PMID: 41128078
Abstract
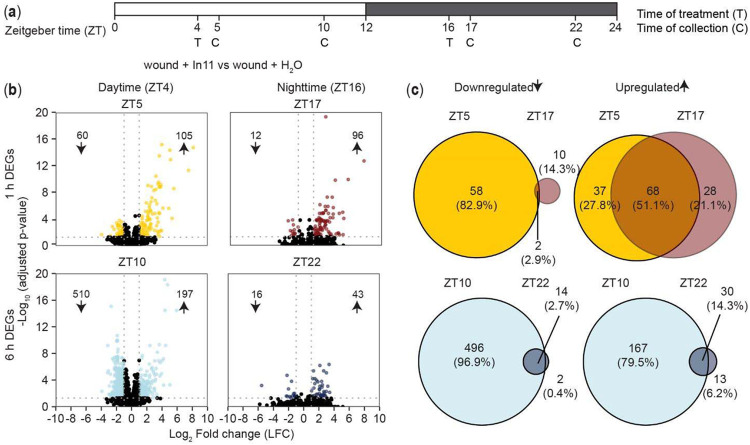
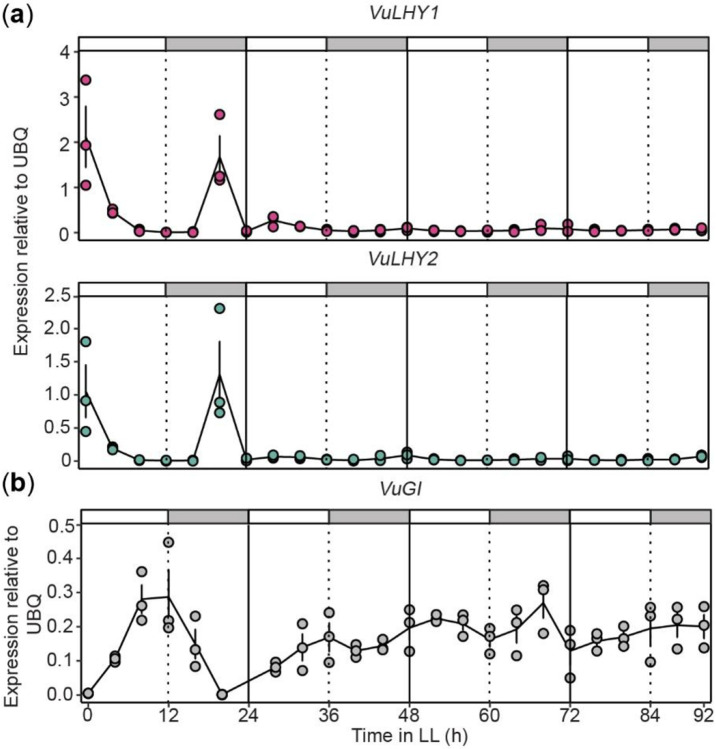
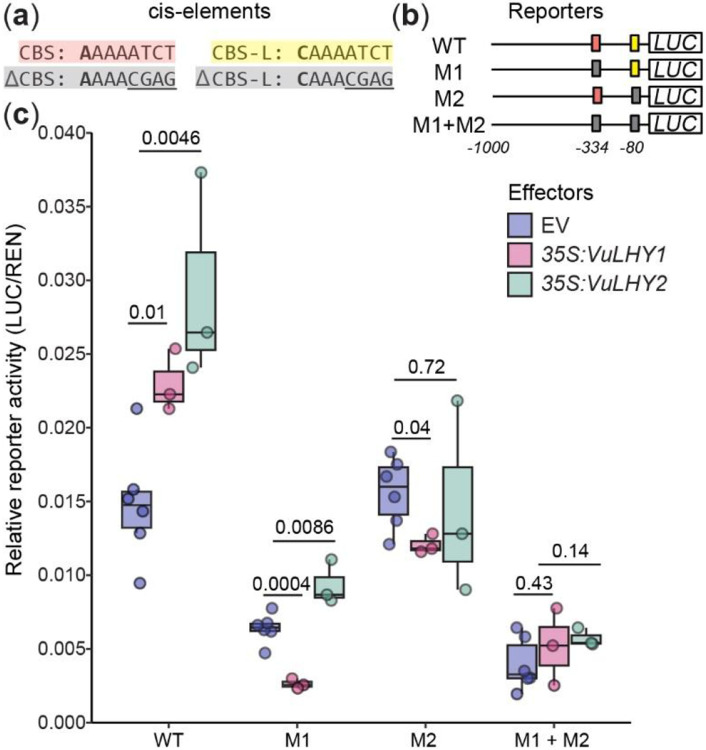
Plants activate induced defenses through the recognition of molecular patterns. Like pathogen-associated molecular patterns (PAMPs), herbivore-associated molecular patterns (HAMPs) can be recognized by cell surface pattern recognition receptors leading to defensive transcriptional changes in host plants. Herbivore-induced defensive outputs are regulated by the circadian clock, but the underlying molecular mechanisms remain unknown. To investigate how the plant circadian clock regulates transcriptional reprogramming of a specific HAMP-induced pathway, we characterized the daytime and nighttime transcriptional response to caterpillar-derived In11 peptide, in the legume crop cowpea (Vigna unguiculata). Using diurnal and free-running conditions, we found that daytime In11 elicitation resulted in stronger late-induced gene expression than nighttime. Plants with a conditional arrhythmic phenotype in constant light (LL) conditions lost time-of-day dependent responses to In11 treatment, and this was associated with arrhythmic expression of circadian clock core transcription factor Late Elongated Hypocotyl VuLHY1 and VuLHY2. Reporter assays with VuLHY homologs indicated that they interact with the promoter of daytime In11-induced Kunitz Trypsin Inhibitor (VuKTI) via a canonical and a polymorphic CCA1/LHY Binding Site (CBS), consistent with a mechanism of direct regulation by circadian clock transcription factors. This study improves our understanding of the time-dependent mechanisms that regulate herbivore-induced gene expression.
Keywords: HAMP; In11; Inceptin; circadian clock; circadian gating; immunity; legumes; time-of-day.
Figures



References
-
- Adams S, Grundy J, Veflingstad SR, Dyer NP, Hannah MA, Ott S, Carré IA. 2018. Circadian control of abscisic acid biosynthesis and signalling pathways revealed by genome-wide analysis of LHY binding targets. The new phytologist 220: 893–907. - PubMed
-
- DeFalco TA, Zipfel C. 2021. Molecular mechanisms of early plant pattern-triggered immune signaling. Molecular cell 81: 3449–3467. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources