

Transcriptional profiling reveals the role of Candida albicans Rap1 in oxidative stress response
- PMID: 39575984
- PMCID: PMC11667096
- DOI: 10.1042/BSR20240689
Transcriptional profiling reveals the role of Candida albicans Rap1 in oxidative stress response
Abstract
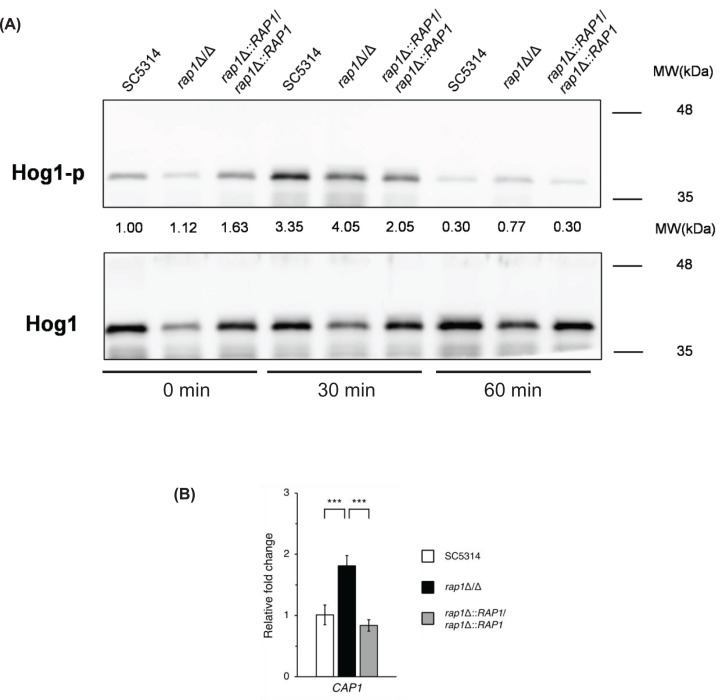
Candida albicans is a member of the human commensal microbiota but can also cause opportunistic infections, including life-threatening invasive candidiasis, particularly in immunocompromised patients. One of the important features of C. albicans commensalism and virulence is its ability to adapt to diverse environmental stress conditions within the host. Rap1 is a DNA-binding protein identified in yeasts, protozoa, and mammalian cells, and it plays multiple functions, including telomere regulation. Intriguingly, our previous study showed that Rap1 is also involved in cell wall integrity, biofilm formation, and virulence in C. albicans. In this work, using RNA-seq analysis and other approaches, the role of C. albicans Rap1 in oxidative stress response was further revealed. The RAP1-deletion mutant exhibited greater resistance to the superoxide generator menadione, a lower level of intracellular reactive oxygen species (ROS) upon menadione treatment, and higher expression levels of superoxide dismutase genes, all in response to oxidative stress. Moreover, the association between Rap1-mediated oxidative stress response and the mitogen-activated protein kinase (MAPK) Hog1, the transcription factor Cap1 and the TOR signalling was also determined. Together, these findings expand our understanding of the complex signalling and transcriptional mechanisms regulating stress responses in C. albicans.
Keywords: Candida albicans; Rap1; oxidative stress.
© 2024 The Author(s).
Conflict of interest statement
The authors declare that there are no competing interests associated with the manuscript.
Figures



Similar articles
-
A small heat shock protein Fmp28 influences virulence by regulating Als3 expression via the cAMP-PKA signaling pathway in Candida albicans.mBio. 2025 Aug 13;16(8):e0125325. doi: 10.1128/mbio.01253-25. Epub 2025 Jun 30. mBio. 2025. PMID: 40586563 Free PMC article.
-
Histone deacetylase Sir2 promotes the systemic Candida albicans infection by facilitating its immune escape via remodeling the cell wall and maintaining the metabolic activity.mBio. 2024 Jun 12;15(6):e0044524. doi: 10.1128/mbio.00445-24. Epub 2024 Apr 29. mBio. 2024. PMID: 38682948 Free PMC article.
-
Transcriptomic insights into Candida albicans adaptation to an anaerobic environment.Microbiol Spectr. 2025 Jul;13(7):e0302424. doi: 10.1128/spectrum.03024-24. Epub 2025 May 22. Microbiol Spectr. 2025. PMID: 40401963 Free PMC article.
-
Candida albicans: Insights into the Biology and Experimental Innovations of a Commonly Isolated Human Fungal Pathogen.ACS Infect Dis. 2025 Jul 11;11(7):1780-1815. doi: 10.1021/acsinfecdis.5c00079. Epub 2025 Jun 13. ACS Infect Dis. 2025. PMID: 40512087 Review.
-
Insights into the evolution of Candidalysin and recent developments.Arch Microbiol. 2025 Jul 31;207(9):206. doi: 10.1007/s00203-025-04414-z. Arch Microbiol. 2025. PMID: 40742540 Review.
Cited by
-
Deletion of RAP1 affects iron homeostasis, azole resistance, and virulence in Candida albicans.mSphere. 2025 May 27;10(5):e0015525. doi: 10.1128/msphere.00155-25. Epub 2025 Apr 23. mSphere. 2025. PMID: 40265929 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous