

BTK regulates microglial function and neuroinflammation in human stem cell models and mouse models of multiple sclerosis
- PMID: 39578444
- PMCID: PMC11584639
- DOI: 10.1038/s41467-024-54430-8
BTK regulates microglial function and neuroinflammation in human stem cell models and mouse models of multiple sclerosis
Abstract
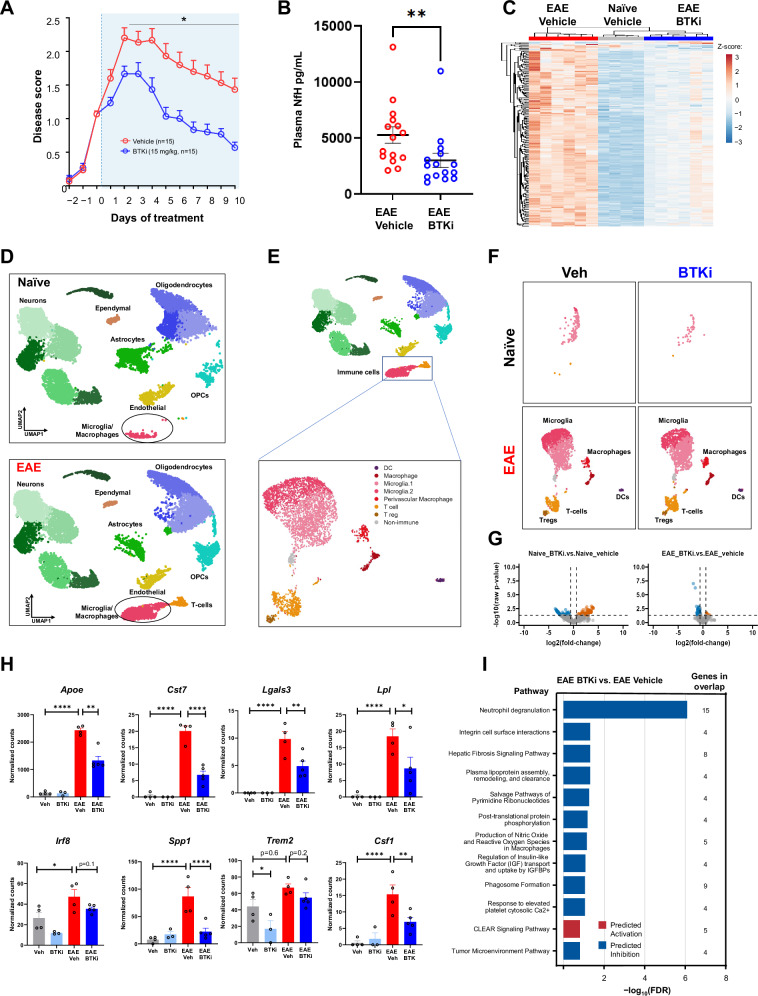
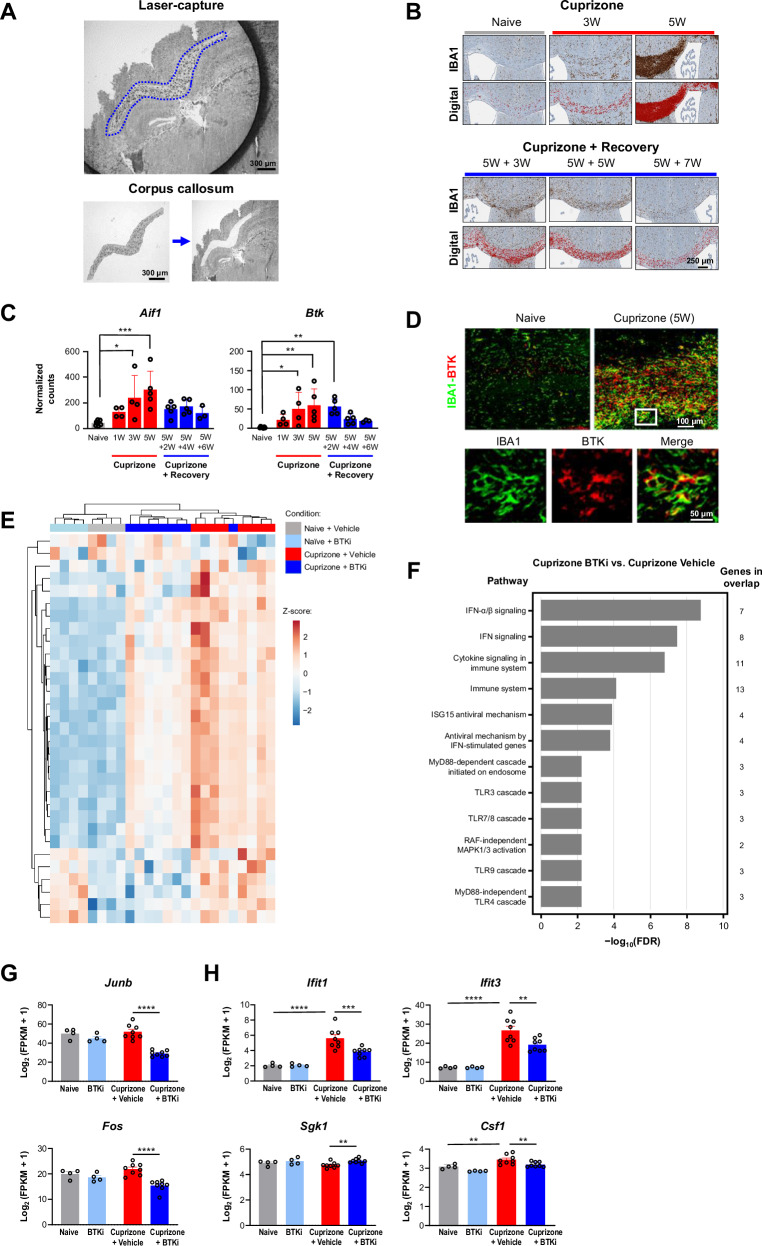
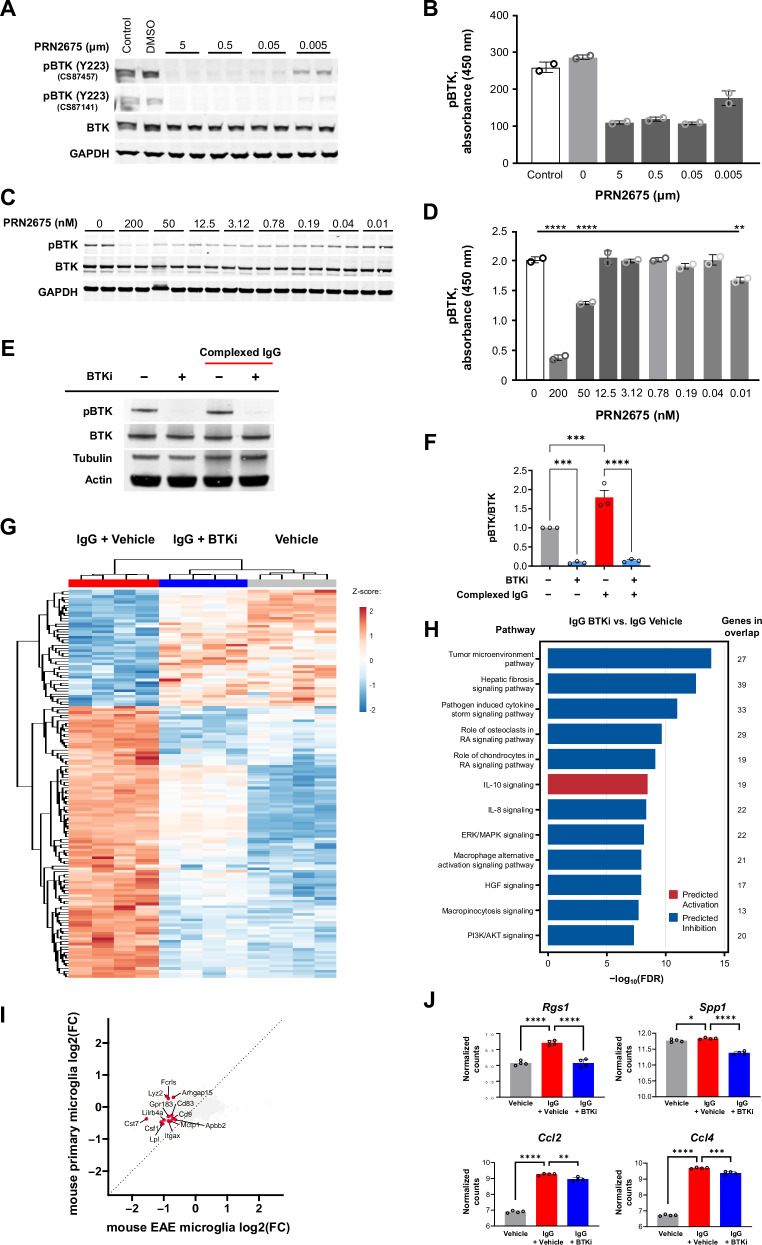
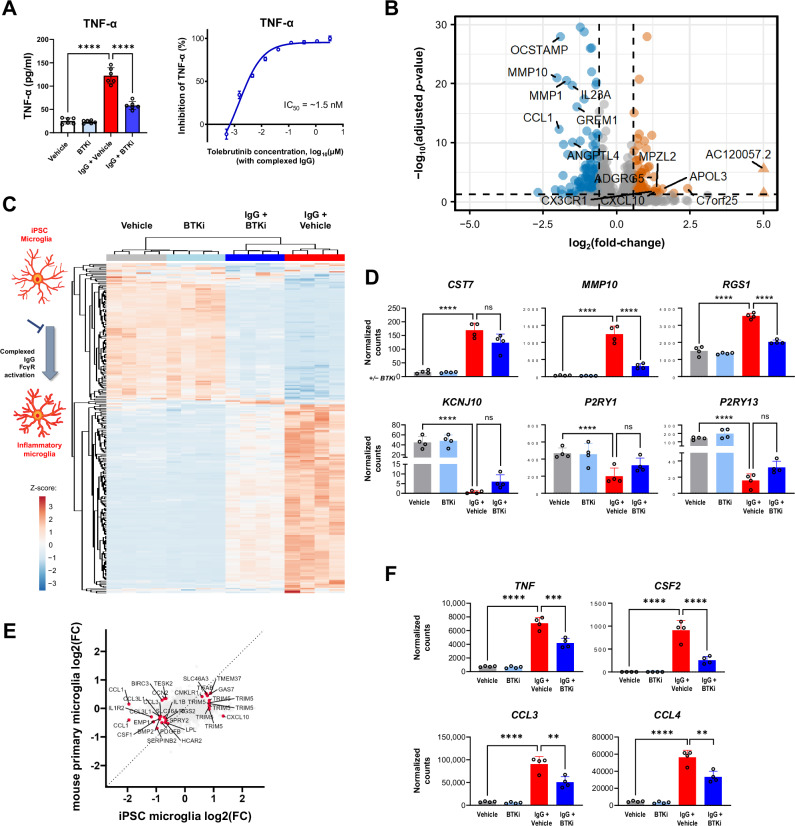
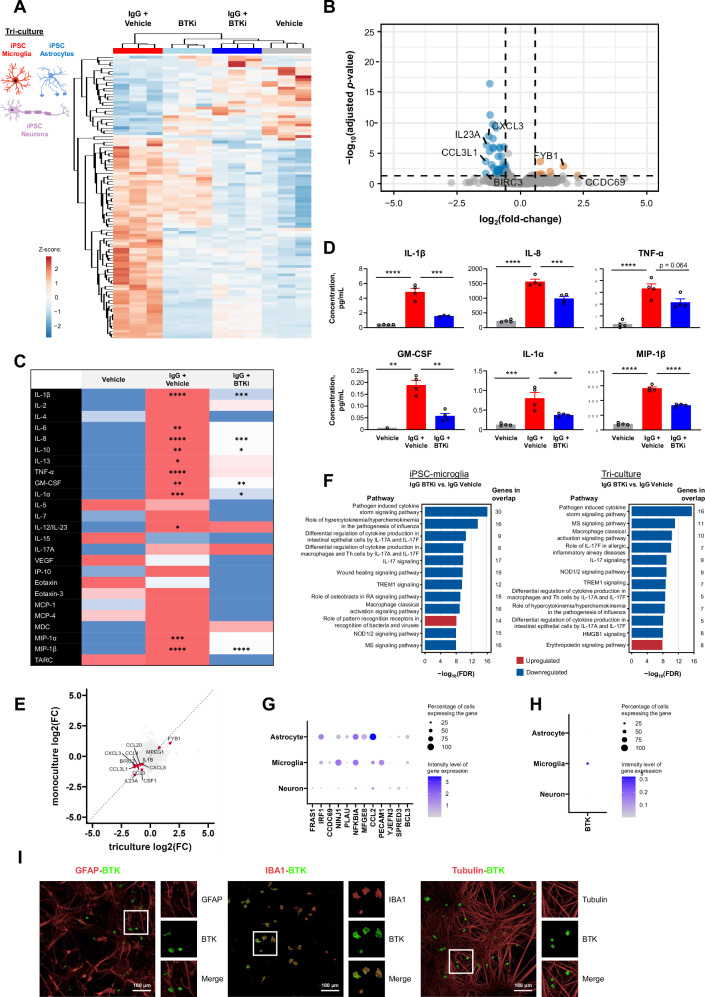
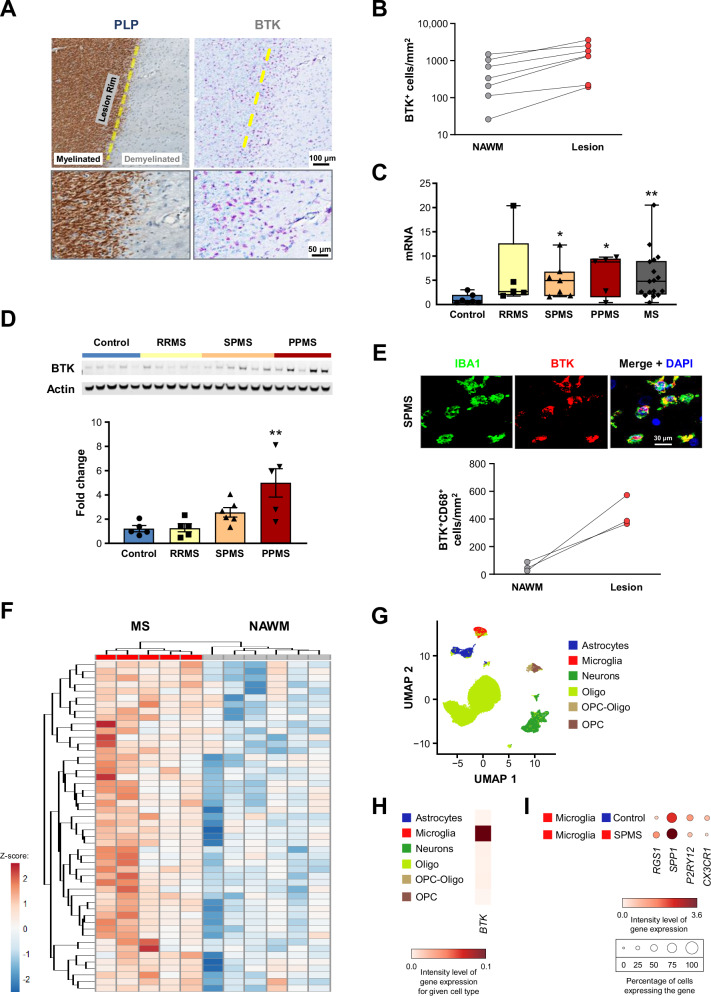
Neuroinflammation in the central nervous system (CNS), driven largely by resident phagocytes, has been proposed as a significant contributor to disability accumulation in multiple sclerosis (MS) but has not been addressed therapeutically. Bruton's tyrosine kinase (BTK) is expressed in both B-lymphocytes and innate immune cells, including microglia, where its role is poorly understood. BTK inhibition may provide therapeutic benefit within the CNS by targeting adaptive and innate immunity-mediated disease progression in MS. Using a CNS-penetrant BTK inhibitor (BTKi), we demonstrate robust in vivo effects in mouse models of MS. We further identify a BTK-dependent transcriptional signature in vitro, using the BTKi tolebrutinib, in mouse microglia, human induced pluripotent stem cell (hiPSC)-derived microglia, and a complex hiPSC-derived tri-culture system composed of neurons, astrocytes, and microglia, revealing modulation of neuroinflammatory pathways relevant to MS. Finally, we demonstrate that in MS tissue BTK is expressed in B-cells and microglia, with increased levels in lesions. Our data provide rationale for targeting BTK in the CNS to diminish neuroinflammation and disability accumulation.
© 2024. The Author(s).
Conflict of interest statement
Competing interests: R.C.G., N.C., and N.P. were employees of Sanofi when this work was undertaken. G.S.W., A.S.B., L.L., M.R.D., N.H., M.L., T.R.H., A.Che, S.R., M.Z., E.H., A.M., T.J.T., and D.O. are employees of Sanofi and may hold shares and/or stock options in the company. ACho and EC declare that they have no competing interests. B.D.T has received consulting fees, speaker honoraria, and/or research funding from Biogen, Disarm Therapeutics, EMD Serono, Novartis, Renovo Neural, and Sanofi; and principal investigator and/or speaking fees from Alkermes, Biogen, Celgene, EMD Serono, Genentech/Roche, Novartis, Sanofi, and TG Therapeutics.
Figures
References
-
- Baecher-Allan, C., Kaskow, B. J. & Weiner, H. L. Multiple sclerosis: mechanisms and immunotherapy. Neuron97, 742–768 (2018). - PubMed
-
- Hauser, S. L. et al. Ofatumumab versus teriflunomide in multiple sclerosis. N. Engl. J. Med.383, 546–557 (2020). - PubMed
-
- Hauser, S. L. et al. Ocrelizumab versus interferon beta-1a in relapsing multiple sclerosis. N. Engl. J. Med.376, 221–234 (2017). - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Medical
