

The PvRBP2b-TfR1 interaction is not essential for reticulocytes invasion by Plasmodium vivax isolates from Cambodia
- PMID: 39578462
- PMCID: PMC11584642
- DOI: 10.1038/s41541-024-01031-7
The PvRBP2b-TfR1 interaction is not essential for reticulocytes invasion by Plasmodium vivax isolates from Cambodia
Abstract
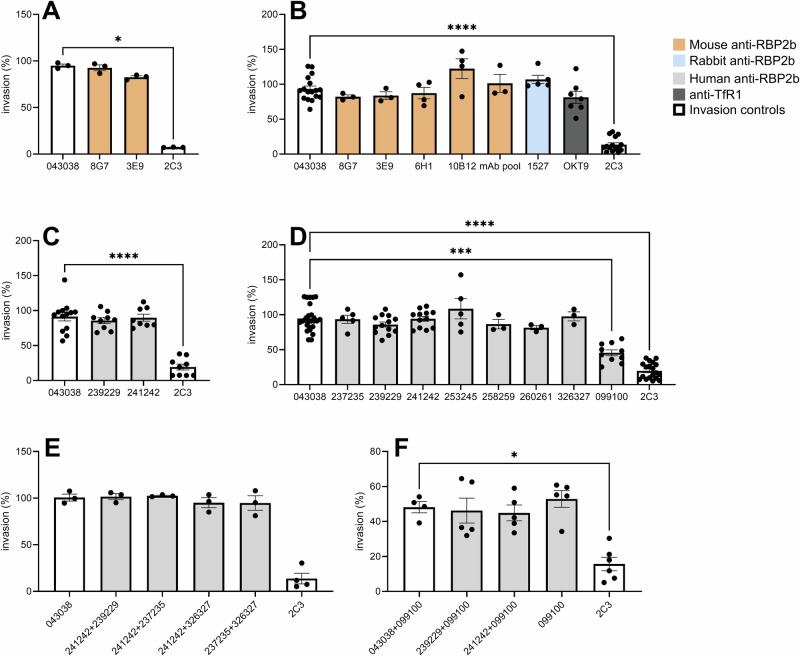
Plasmodium vivax is the most widespread of the different Plasmodium species able to infect humans and is responsible for most malaria cases outside Africa. An effective, strain-transcending vaccine that alleviates or suppresses erythrocyte invasion would be a game-changer in eliminating vivax malaria. Recently, the binding of P. vivax Reticulocyte Binding Protein 2b (PvRBP2b) to human Transferrin receptor (TfR1) has been described as essential for reticulocyte invasion, making this parasite protein an appealing vaccine candidate. Here, using P. vivax Cambodian clinical isolates in robust ex vivo invasion assays, we show that anti-PvRBP2b polyclonal and monoclonal antibodies that inhibit binding of PvRBP2b to TfR1 do not block P. vivax invasion into reticulocytes even at high concentrations. Anti-TfR1 antibodies do not inhibit P. vivax invasion either. Combinations at high concentrations of human monoclonal antibodies targeting different PvRBP2b epitopes do not inhibit invasion. Combinations of anti-PvRBP2b with anti-PvDBP do not enhance invasion inhibition caused by anti-PvDBP alone. We also show that the invasion of Cambodian P. vivax is trypsin-resistant while TfR1 is trypsin-sensitive, and we demonstrate that TfR1 is not recycled following trypsin treatment. We determined the PvRBP2b sequence of all isolates used in the invasion assays and analyzed polymorphism within epitopes recognized by anti-PvRBP2b antibodies. We show that polymorphism does not explain the absence of neutralization. Anti-PvRBP2b polyclonal antibodies recognized all four isolates tested in immunofluorescence assays while not inhibiting P. vivax invasion. Overall, our results demonstrate that PvRBP2b binding to TfR1 is not essential for invasion into reticulocytes of P. vivax Cambodian strains questioning the relevance of PvRBP2b as vaccine candidate.
© 2024. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures



References
-
- Mueller, I. et al. Key gaps in the knowledge of Plasmodium vivax, a neglected human malaria parasite. Lancet Infect. Dis.9, 555–566 (2009). - PubMed
Grants and funding
- R01AI173171/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
- R01 AI173171/AI/NIAID NIH HHS/United States
- R01 AI175134/AI/NIAID NIH HHS/United States
- R01AI162947/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
- R01 AI162947/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
