

Ileal microbial microbiome and its secondary bile acids modulate susceptibility to nonalcoholic steatohepatitis in dairy goats
- PMID: 39578870
- PMCID: PMC11585128
- DOI: 10.1186/s40168-024-01964-0
Ileal microbial microbiome and its secondary bile acids modulate susceptibility to nonalcoholic steatohepatitis in dairy goats
Abstract
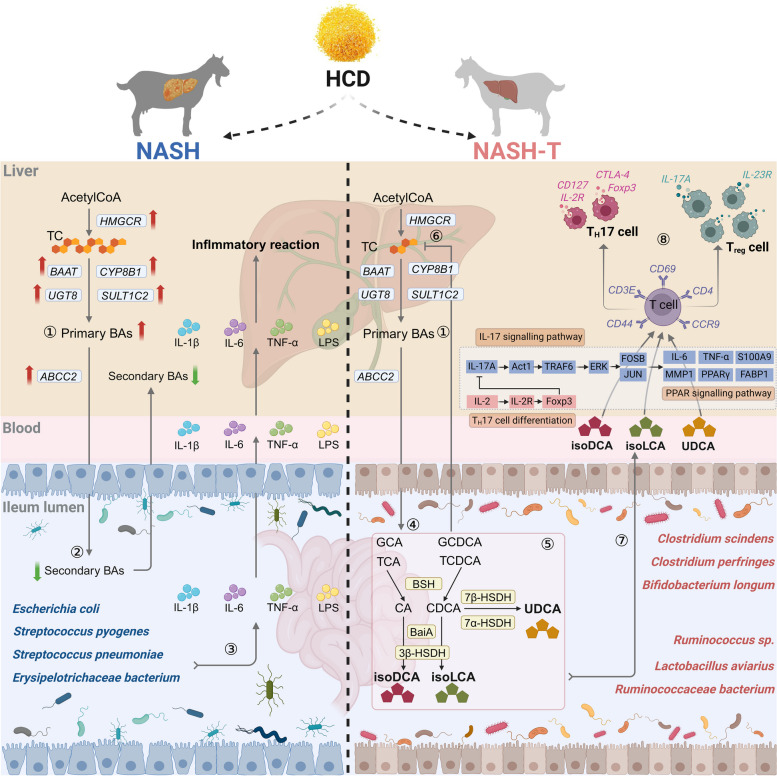
Background: Liver damage from nonalcoholic steatohepatitis (NASH) presents a significant challenge to the health and productivity of ruminants. However, the regulatory mechanisms behind variations in NASH susceptibility remain unclear. The gut‒liver axis, particularly the enterohepatic circulation of bile acids (BAs), plays a crucial role in regulating the liver diseases. Since the ileum is the primary site for BAs reabsorption and return to the liver, we analysed the ileal metagenome and metabolome, liver and serum metabolome, and liver single-nuclei transcriptome of NASH-resistant and susceptible goats together with a mice validation model to explore how ileal microbial BAs metabolism affects liver metabolism and immunity, uncovering the key mechanisms behind varied NASH pathogenesis in dairy goats.
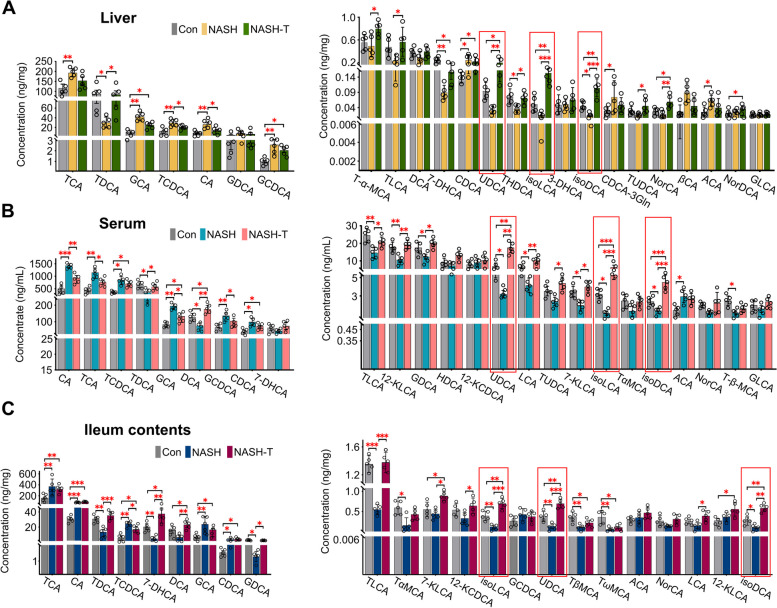
Results: In NASH goats, increased total cholesterol (TC), triglyceride (TG), and primary BAs and decreased secondary BAs in the liver and serum promoted hepatic fat accumulation. Increased ileal Escherichia coli, Erysipelotrichaceae bacterium and Streptococcus pneumoniae as well as proinflammatory compounds damaged ileal histological morphology, and increased ileal permeability contributes to liver inflammation. In NASH-tolerance (NASH-T) goats, increased ursodeoxycholic acid (UDCA), isodeoxycholic acid (isoDCA) and isolithocholic acid (isoLCA) in the liver, serum and ileal contents were attributed to ileal secondary BAs-producing bacteria (Clostridium, Bifidobacterium and Lactobacillus) and key microbial genes encoding enzymes. Meanwhile, decreased T-helper 17 (TH17) cells and increased regulatory T (Treg) cells proportion were identified in both liver and ileum of NASH-T goats. To further validate whether these key BAs affected the progression of NASH by regulating the proliferation of TH17 and Treg cells, the oral administration of bacterial UDCA, isoDCA and isoLCA to a high-fat diet-induced NASH mouse model confirmed the amelioration of NASH through the TH17 cell differentiation/IL-17 signalling/PPAR signalling pathway by these bacterial secondary BAs.
Conclusion: This study revealed the roles of ileal microbiome and its secondary BAs in resilience and susceptibility to NASH by affecting the hepatic Treg and TH17 cells proportion in dairy goats. Bacterial UDCA, isoDCA and isoLCA were demonstrated to alleviate NASH and could be novel postbiotics to modulate and improve the liver health in ruminants. Video Abstract.
Keywords: Dairy goats; Ileal microbiome; Intrahepatic TH17 cells and Treg cells; Nonalcoholic steatohepatitis susceptibility; Secondary bile acids.
© 2024. The Author(s).
Conflict of interest statement
Declarations. Ethics approval and consent to participate: All the experimental designs and protocols used in the present study were approved by the Institutional Animal Care and Use Committee (IACUC) of Northwest A&F University (Shaanxi, China, approval number: NWAFU-DK-2022020) and were in accordance with the recommendations of the university’s guidelines for animal research. Consent for publication: Not applicable. Competing interests: The authors declare no competing interests.
Figures





References
-
- Soares RAN, Vargas G, Muniz MMM, Soares MAM, Cánovas A, Schenkel F, Squires EJ. Differential gene expression in dairy cows under negative energy balance and ketosis: a systematic review and meta-analysis. J Dairy Sci. 2021;104(1):602–15. - PubMed
-
- Chirivi M, Cortes-Beltran D, Munsterman A, O’Connor A, Contreras GA. Lipolysis inhibition as a treatment of clinical ketosis in dairy cows: a randomized clinical trial. J Dairy Sci. 2023;106(12):9514–31. - PubMed
-
- Du X, Chen M, Fang Z, Shao Q, Yu H, Hao X, Gao X, Ju L, Li C, Yang Y, Song Y, Lei L, Liu G, Li X. Evaluation of hepatic AMPK, mTORC1, and autophagy-lysosomal pathway in cows with mild or moderate fatty liver. J Dairy Sci. 2024;107(5):3269–79. - PubMed
-
- Chandra Roy A, Wang Y, Zhang H, Roy S, Dai H, Chang G, Shen X. Sodium butyrate mitigates iE-DAP induced inflammation caused by high-concentrate feeding in liver of dairy goats. J Agric Food Chem. 2018;66(34):8999–9009. - PubMed
-
- Zhang H, Shi H, Xie W, Meng M, Wang Y, Ma N, Chang G, Shen X. Subacute ruminal acidosis induces pyroptosis via the mitophagy-mediated NLRP3 inflammasome activation in the livers of dairy cows fed a high-grain diet. J Dairy Sci. 2024;24(S0022-0302):00042. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
