

Single-cell transcriptomics unveils molecular signatures of neuronal vulnerability in a mouse model of prion disease that overlap with Alzheimer's disease
- PMID: 39580485
- PMCID: PMC11585576
- DOI: 10.1038/s41467-024-54579-2
Single-cell transcriptomics unveils molecular signatures of neuronal vulnerability in a mouse model of prion disease that overlap with Alzheimer's disease
Abstract
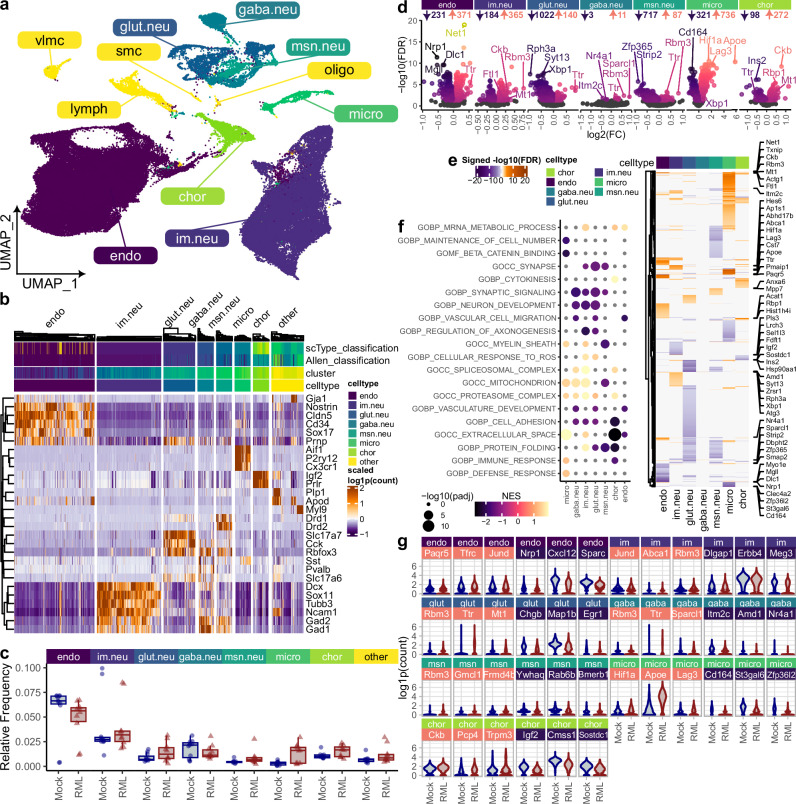
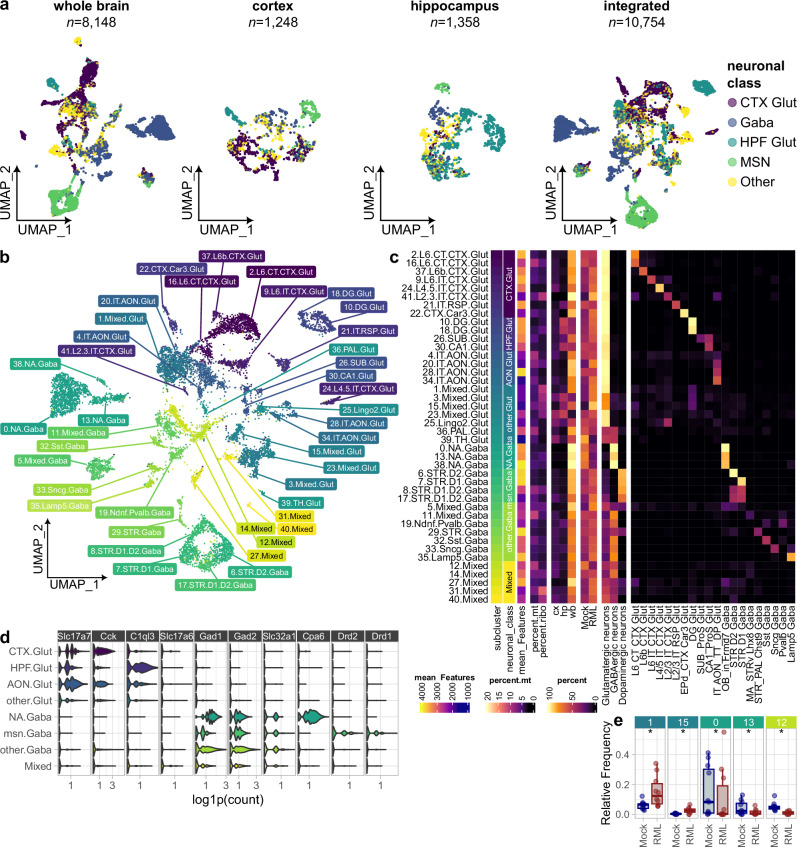
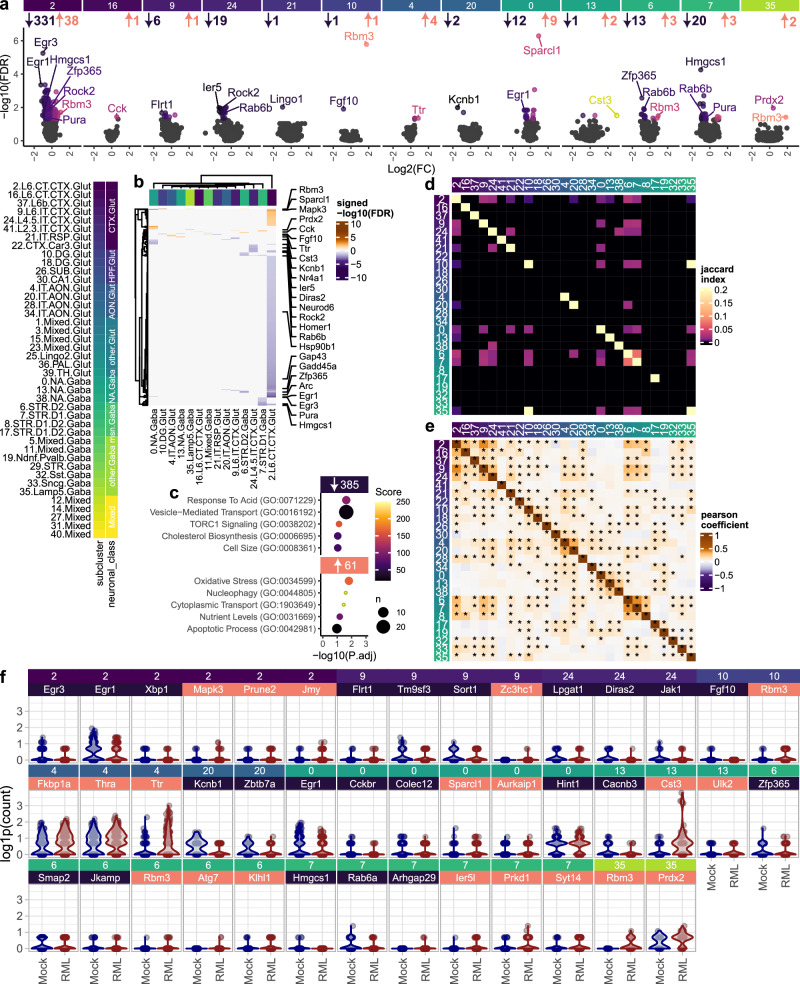
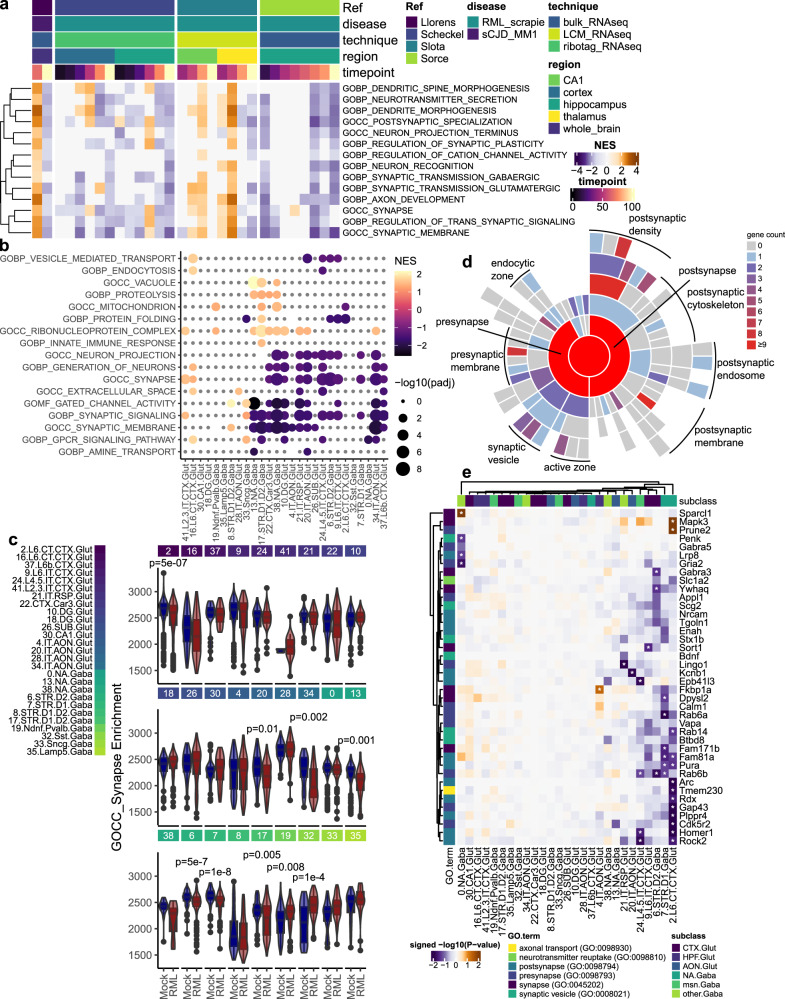
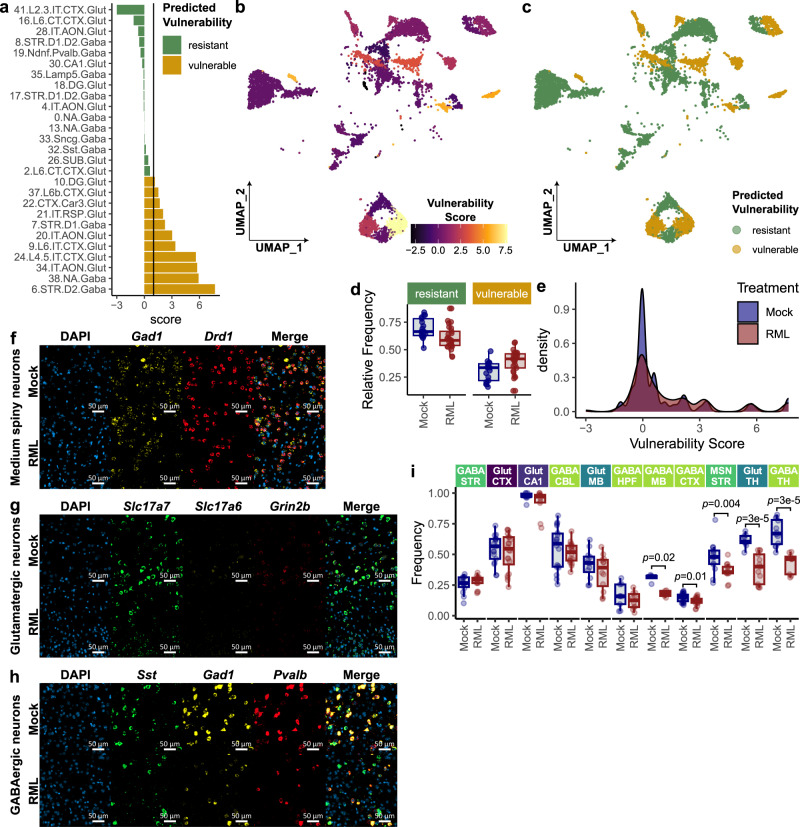
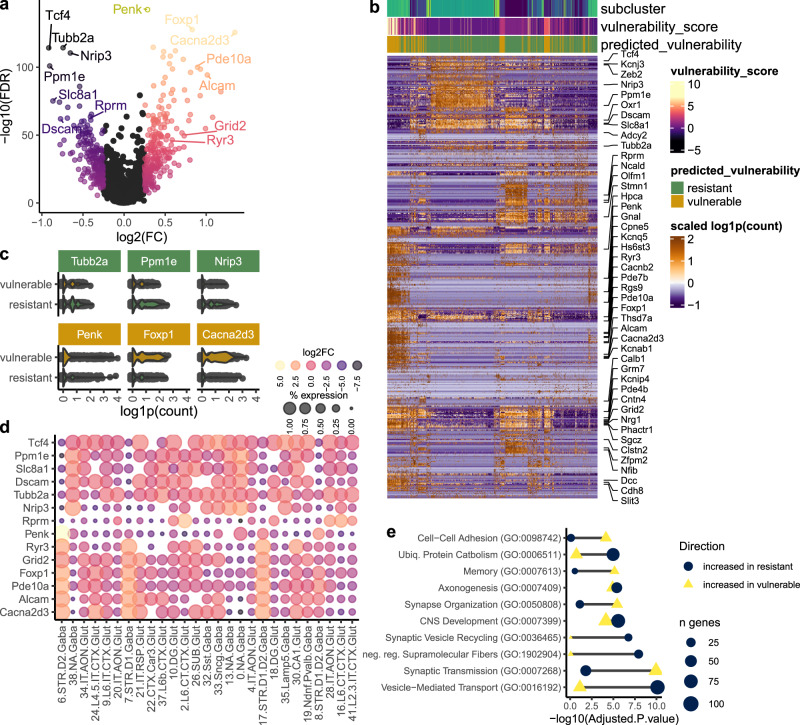
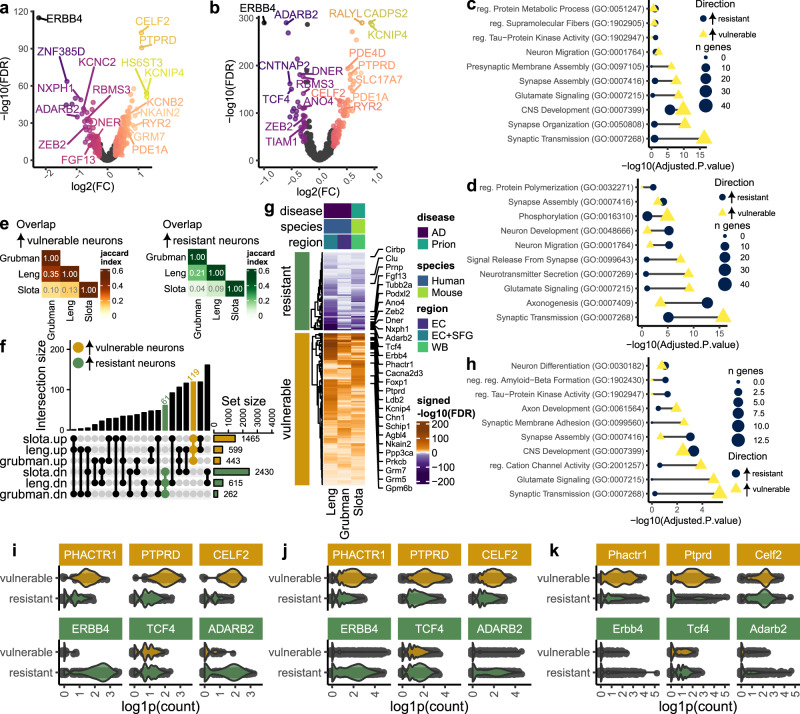
Understanding why certain neurons are more sensitive to dysfunction and death caused by misfolded proteins could provide therapeutically relevant insights into neurodegenerative disorders. Here, we harnessed single-cell transcriptomics to examine live neurons isolated from prion-infected female mice, aiming to identify and characterize prion-vulnerable neuronal subsets. Our analysis revealed distinct transcriptional responses across neuronal subsets, with a consistent pathway-level depletion of synaptic gene expression in damage-vulnerable neurons. By scoring neuronal damage based on the magnitude of depleted synaptic gene expression, we identified a diverse spectrum of prion-vulnerable glutamatergic, GABAergic, and medium spiny neurons. Comparison between prion-vulnerable and resistant neurons highlighted baseline gene expression differences that could influence neuronal vulnerability. For instance, the neuroprotective cold-shock protein Rbm3 exhibited higher baseline gene expression in prion-resistant neurons and was robustly upregulated across diverse neuronal classes upon prion infection. We also identified vulnerability-correlated transcripts that overlapped between prion and Alzheimer's disease. Our findings not only demonstrate the potential of single-cell transcriptomics to identify damage-vulnerable neurons, but also provide molecular insights into neuronal vulnerability and highlight commonalties across neurodegenerative disorders.
© 2024. Crown.
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures

References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
