

Modulating the immunodominance hierarchy of immunoglobulin germline-encoded structural motifs targeting the influenza hemagglutinin stem
- PMID: 39580804
- PMCID: PMC11672684
- DOI: 10.1016/j.celrep.2024.114990
Modulating the immunodominance hierarchy of immunoglobulin germline-encoded structural motifs targeting the influenza hemagglutinin stem
Abstract
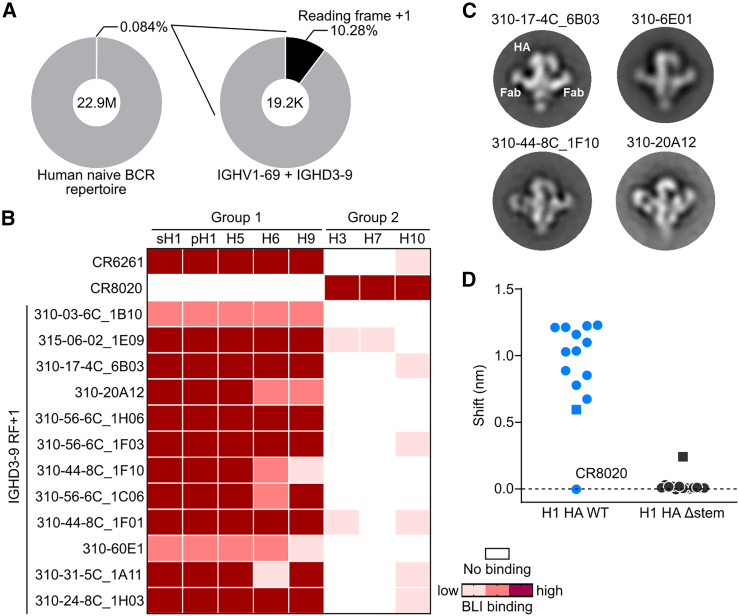
Antibodies targeting epitopes through germline-encoded motifs can be found in different individuals. While these public antibodies are often beneficial, they also pose hurdles for subdominant antibodies to emerge. Here, we use transgenic mice that reproduce the human IGHV1-69∗01 germline-encoded antibody response to the conserved stem epitope on group 1 hemagglutinin (HA) of influenza A virus to show that this germline-endowed response can be overridden by a subdominant yet cross-group reactive public antibody response. Immunization with a non-cognate group 2 HA stem enriched B cells harboring the IGHD3-9 gene, thereby switching from IGHV1-69- to IGHD3-9-encoded motif-dependent epitope recognition. These IGHD3-9 antibodies bound, neutralized, and conferred cross-group protection in mice against influenza A viruses. A cryoelectron microscopy (cryo-EM) structure of an IGHD3-9 antibody resembled the human broadly neutralizing antibody FI6v3, which uses IGHD3-9. Together, our findings offer insights into vaccine regimens that engage an immunoglobulin repertoire with broader cross-reactivity to influenza A viruses.
Keywords: B cell; CP: Immunology; HA stem; IGHD3-9; IGHV1-69; bnAb; broadly neutralizing antibody; germline-targeting; hemagglutinin; influenza virus; nanoparticle.
Published by Elsevier Inc.
Conflict of interest statement
Declaration of interests S.M.M., B.S.G., and M.K. are listed as inventors of patents and patent applications on vaccine immunogens used in this study filed by the NIH, US Department of Health and Human Services.
Figures






References
-
- Iuliano A.D., Roguski K.M., Chang H.H., Muscatello D.J., Palekar R., Tempia S., Cohen C., Gran J.M., Schanzer D., Cowling B.J., et al. Estimates of global seasonal influenza-associated respiratory mortality: a modelling study. Lancet. 2018;391:1285–1300. doi: 10.1016/S0140-6736(17)33293-2. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
