

N-acetylaspartate mitigates pro-inflammatory responses in microglial cells by intersecting lipid metabolism and acetylation processes
- PMID: 39587614
- PMCID: PMC11587775
- DOI: 10.1186/s12964-024-01947-6
N-acetylaspartate mitigates pro-inflammatory responses in microglial cells by intersecting lipid metabolism and acetylation processes
Abstract
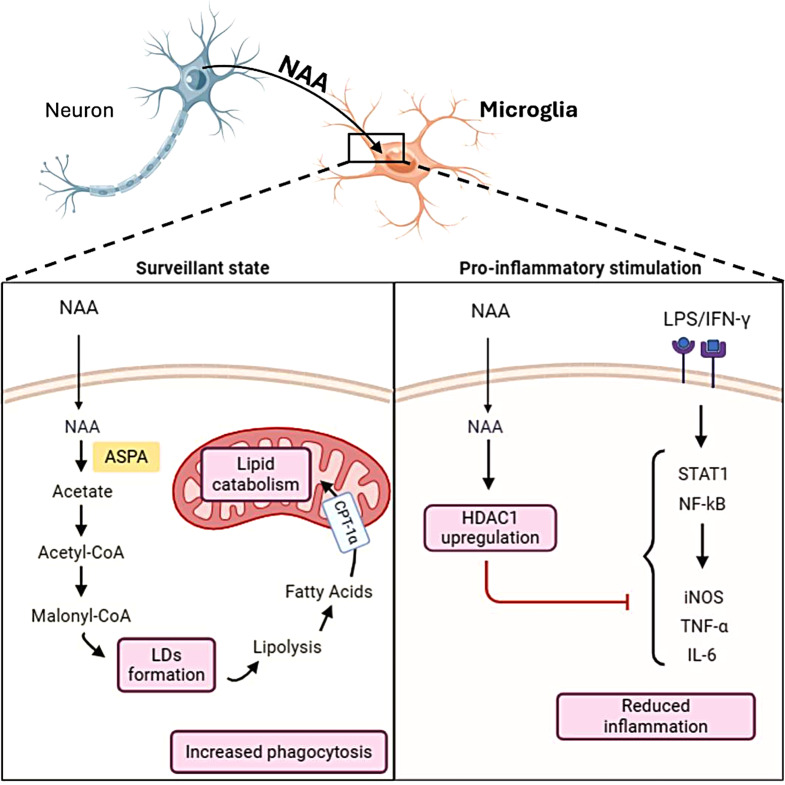
Background: Microglia play a crucial role in brain development and repair by facilitating processes such as synaptic pruning and debris clearance. They can be activated in response to various stimuli, leading to either pro-inflammatory or anti-inflammatory responses associated with specific metabolic alterations. The imbalances between microglia activation states contribute to chronic neuroinflammation, a hallmark of neurodegenerative diseases. N-acetylaspartate (NAA) is a brain metabolite predominantly produced by neurons and is crucial for central nervous system health. Alterations in NAA metabolism are observed in disorders such as Multiple Sclerosis and Canavan disease. While NAA's role in oligodendrocytes and astrocytes has been investigated, its impact on microglial function remains less understood.
Methods: The murine BV2 microglial cell line and primary microglia were used as experimental models. Cells were treated with exogenous NAA and stimulated with LPS/IFN-γ to reproduce the pro-inflammatory phenomenon. HPLC and immunofluorescence analysis were used to study lipid metabolism following NAA treatment. Automated fluorescence microscopy was used to analyze phagocytic activity. The effects on the pro-inflammatory response were evaluated by analysis of protein/mRNA expression and ChIP assay of typical inflammatory markers.
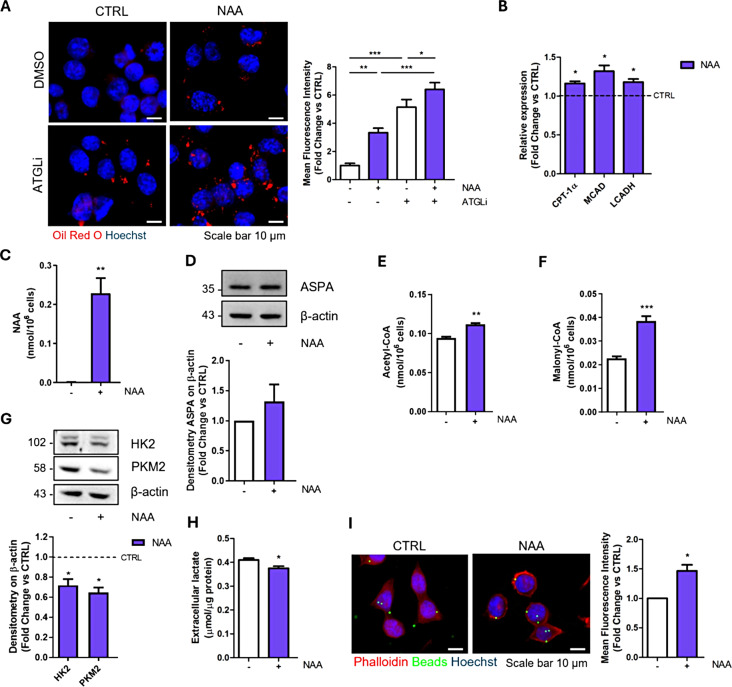
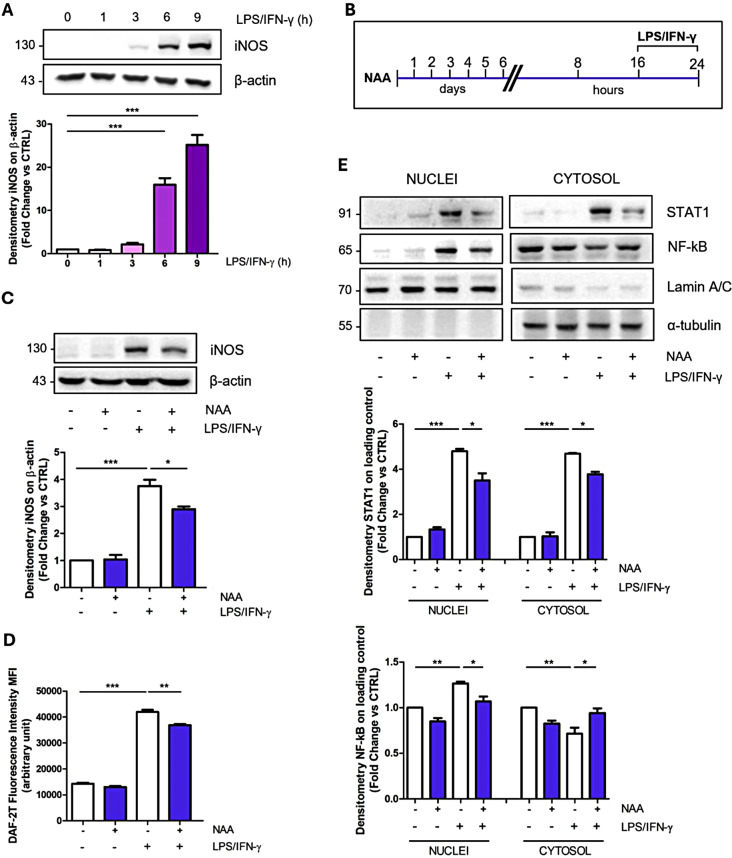
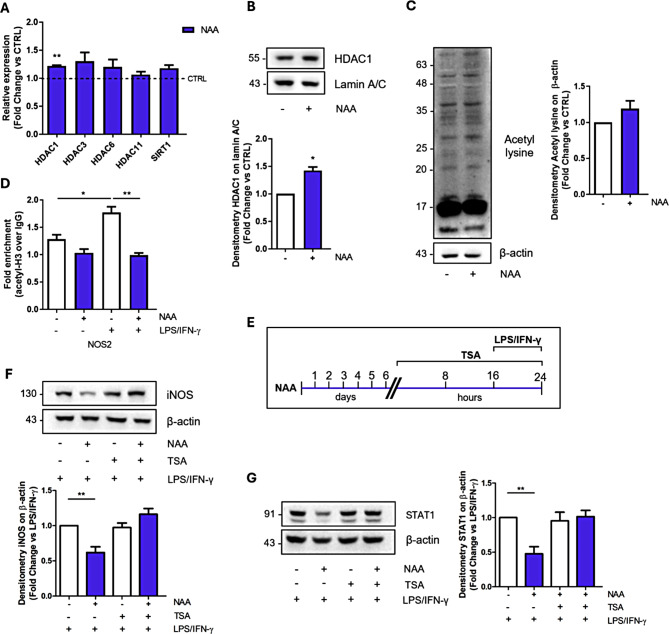
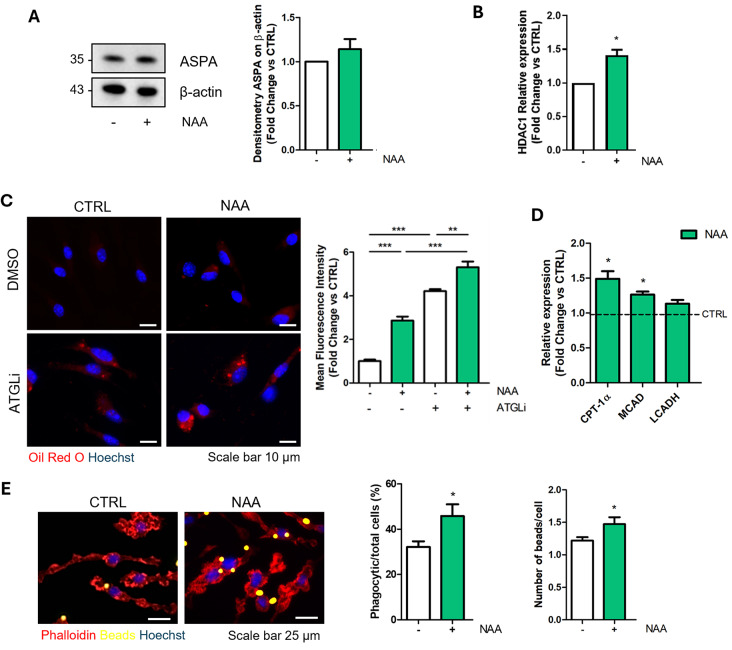
Results: NAA treatment promotes an increase in both lipid synthesis and degradation, and enhances the phagocytic activity of BV2 cells, thus fostering surveillant microglia characteristics. Importantly, NAA decreases the pro-inflammatory state induced by LPS/IFN-γ via the activation of histone deacetylases (HDACs). These findings were validated in primary microglial cells, highlighting the impact on cellular metabolism and inflammatory responses.
Conclusions: The study highlighted the role of NAA in reinforcing the oxidative metabolism of surveillant microglial cells and, most importantly, in buffering the inflammatory processes characterizing reactive microglia. These results suggest that the decreased levels of NAA observed in neurodegenerative disorders can contribute to chronic neuroinflammation.
Keywords: Anti-inflammatory response; Histone deacetylases; Lipid turnover; Microglial polarization; NAA; Oxidative metabolism.
© 2024. The Author(s).
Conflict of interest statement
Declarations. Ethics approval and consent to participate: Not applicable. Consent for publication: All authors are aware of their work and approve of the content of the article and the fact that they are listed as authors of the article. Competing interests: The authors declare no competing interests.
Figures

References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
