

Chemical activation of ABA signaling in grapevine through the iSB09 and AMF4 ABA receptor agonists enhances water use efficiency
- PMID: 39588706
- PMCID: PMC11590044
- DOI: 10.1111/ppl.14635
Chemical activation of ABA signaling in grapevine through the iSB09 and AMF4 ABA receptor agonists enhances water use efficiency
Abstract
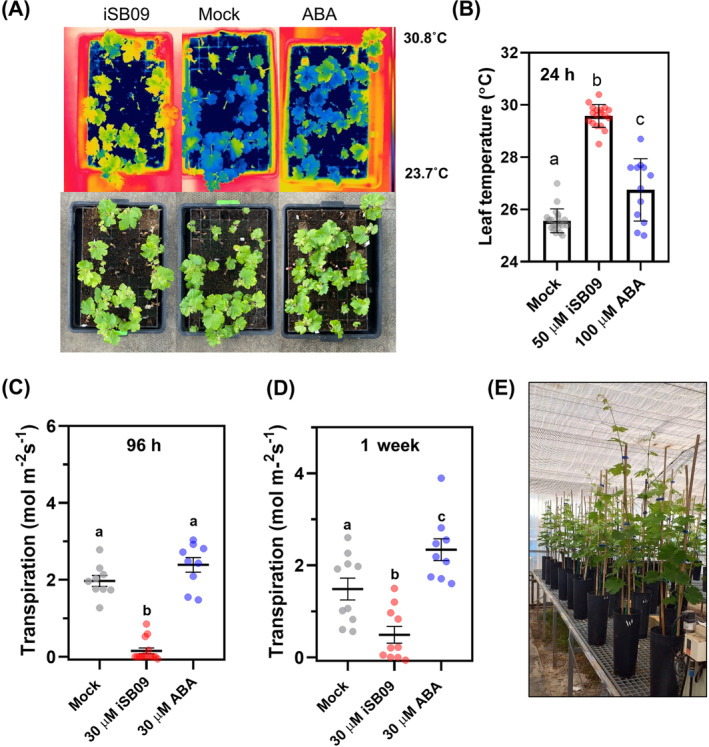
Grapevine (Vitis vinifera L.) is the world's third most valuable horticultural crop, and the current environmental scenario is massively shifting the grape cultivation landscape. The increase in heatwaves and drought episodes alter fruit ripening, compromise grape yield and vine survival, intensifying the pressure on using limited water resources. ABA is a key phytohormone that reduces canopy transpiration and helps plants to cope with water deficit. However, the exogenous application of ABA is impractical because it suffers fast catabolism, and UV-induced isomerization abolishes its bioactivity. Consequently, there is an emerging field for developing molecules that act as ABA receptor agonists and modulate ABA signaling but have a longer half-life. We have explored the foliar application of the iSB09 and AMF4 agonists in the two grapevine cultivars cv. 'Bobal' and 'Tempranillo' to induce an ABA-like response to facilitate plant adaptation to drought. The results indicate that iSB09 and AMF4 act through the VviPYL1-like, VviPYL4-like, and VviPYL8-like ABA receptors to trigger stomatal closure, reduce plant transpiration, and increase water use efficiency. Structural and bioinformatic analysis of VviPYL1 in complex with ABA or these agonists revealed key structural determinants for efficient ligand binding, providing a mechanistic framework to understand receptor activation by the ligands. Physiological analyses further demonstrated that iSB09 has a more sustained effect on reducing transpiration than ABA, and agonist spraying of grapevine leaves protected PSII during drought stress. These findings offer innovative approaches to strengthen the vine's response to water stress and reduce plant consumptive water use under limited soil water conditions.
© 2024 The Author(s). Physiologia Plantarum published by John Wiley & Sons Ltd on behalf of Scandinavian Plant Physiology Society.
Figures






References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
