

Comparison of Microalgae Nannochloropsis oceanica and Chlorococcum amblystomatis Lipid Extracts Effects on UVA-Induced Changes in Human Skin Fibroblasts Proteome
- PMID: 39590789
- PMCID: PMC11595653
- DOI: 10.3390/md22110509
Comparison of Microalgae Nannochloropsis oceanica and Chlorococcum amblystomatis Lipid Extracts Effects on UVA-Induced Changes in Human Skin Fibroblasts Proteome
Abstract
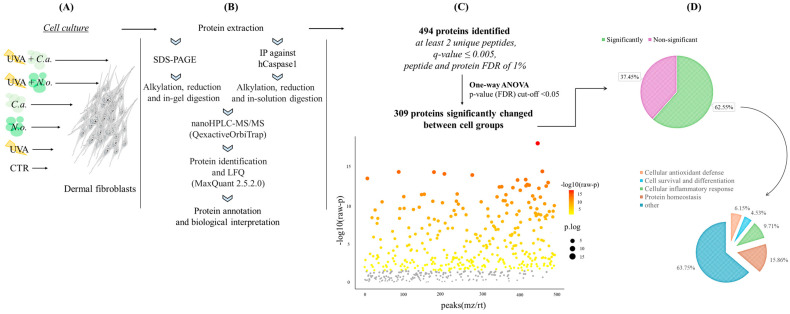
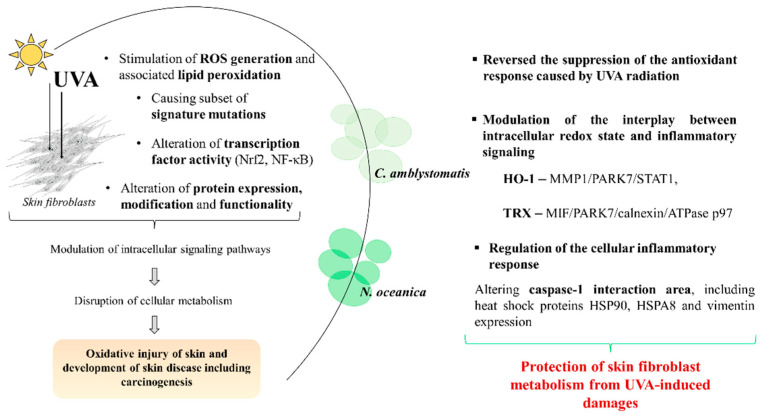
Lipid extracts from the microalgae Nannochloropsis oceanica and Chlorococcum amblystomatis have great potential to prevent ultraviolet A (UVA)-induced metabolic disorders. Therefore, the aim of this study has been to analyze their cytoprotective effect, focused on maintaining intracellular redox balance and inflammation in UVA-irradiated skin fibroblasts, at the proteome level. The above lipid extracts reversed the suppression of the antioxidant response caused by UVA radiation, which was more visible in the case of C. amblystomatis. Modulations of interactions between heme oxygenase-1 and matrix metalloproteinase 1/Parkinson's disease protein 7/transcript1-α/β, as well as thioredoxin and migration inhibitory factor/Parkinson's disease protein 7/calnexin/ATPase p97, created key molecular signaling underlying their cytoprotective actions. Moreover, they reduced pro-inflammatory processes in the control group but they also showed the potential to regulate the cellular inflammatory response by changing inflammasome signaling associated with the changes in the caspase-1 interaction area, including heat shock proteins HSP90, HSPA8, and vimentin. Therefore, lipid extracts from N. oceanica and C. amblystomatis protect skin fibroblast metabolism from UVA-induced damage by restoring the redox balance and regulating inflammatory signaling pathways. Thus, those extracts have proven to have great potential to be used in cosmetic or cosmeceutical products to protect the skin against the effects of solar radiation. However, the possibility of their use requires the evaluation of their effects at the skin level in in vivo and clinical studies.
Keywords: Chlorococcum amblystomatis; Nannochloropsis oceanica; UVA radiation; cytoprotective effect; fibroblast; inflammation; lipid extracts; oxidative stress; proteomics.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures





References
-
- Zhao H.-C., Xiao T., Chen Y.-J. Ultraviolet Induced Skin Inflammation. Int. J. Dermatol. Venereol. 2021;4:229–235. doi: 10.1097/JD9.0000000000000144. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
