

B and T Cell Bi-Cistronic Multiepitopic Vaccine Induces Broad Immunogenicity and Provides Protection Against SARS-CoV-2
- PMID: 39591118
- PMCID: PMC11598604
- DOI: 10.3390/vaccines12111213
B and T Cell Bi-Cistronic Multiepitopic Vaccine Induces Broad Immunogenicity and Provides Protection Against SARS-CoV-2
Abstract
Background: The COVID-19 pandemic, caused by SARS-CoV-2, has highlighted the need for vaccines targeting both neutralizing antibodies (NAbs) and long-lasting cross-reactive T cells covering multiple viral proteins to provide broad and durable protection against emerging variants.
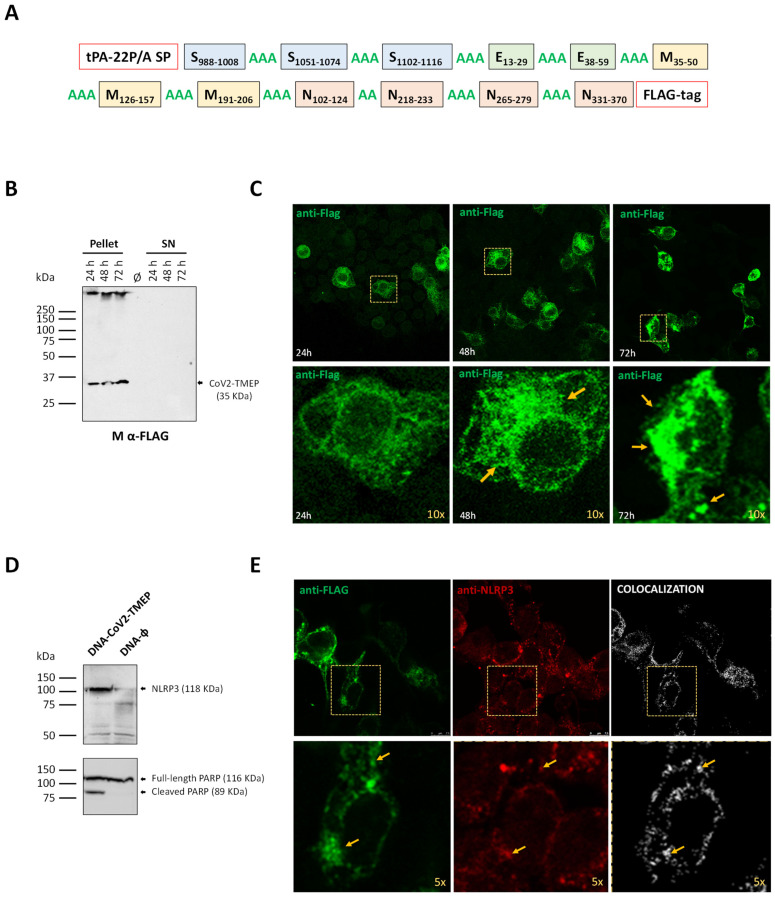
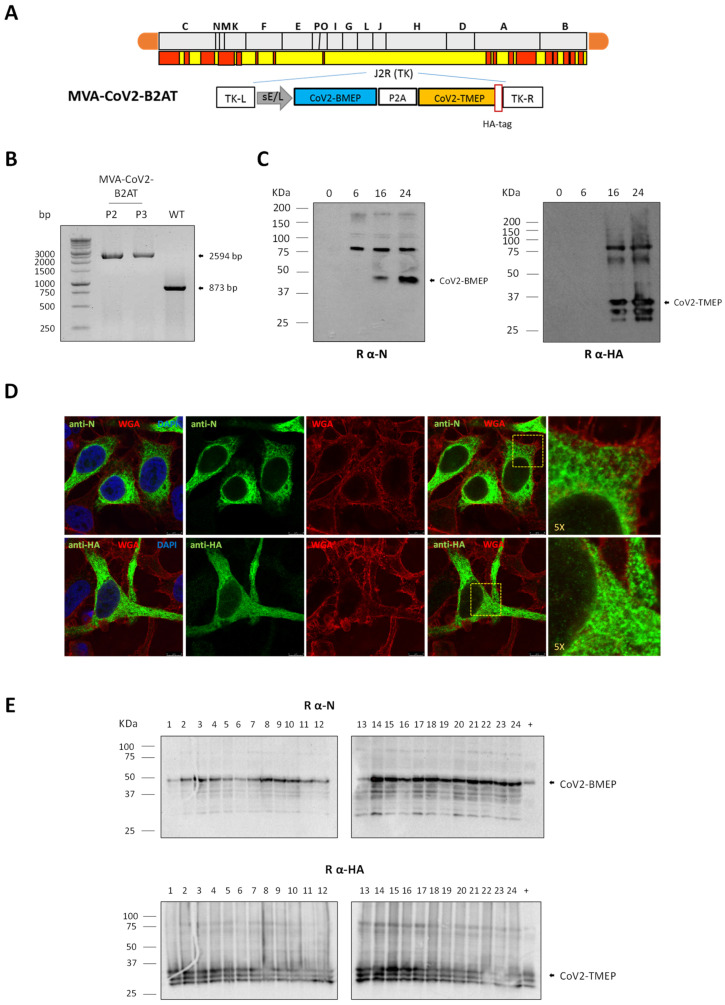
Methods: To address this, here we developed two vaccine candidates, namely (i) DNA-CoV2-TMEP, expressing the multiepitopic CoV2-TMEP protein containing immunodominant and conserved T cell regions from SARS-CoV-2 structural proteins, and (ii) MVA-CoV2-B2AT, encoding a bi-cistronic multiepitopic construct that combines conserved B and T cell overlapping regions from SARS-CoV-2 structural proteins.
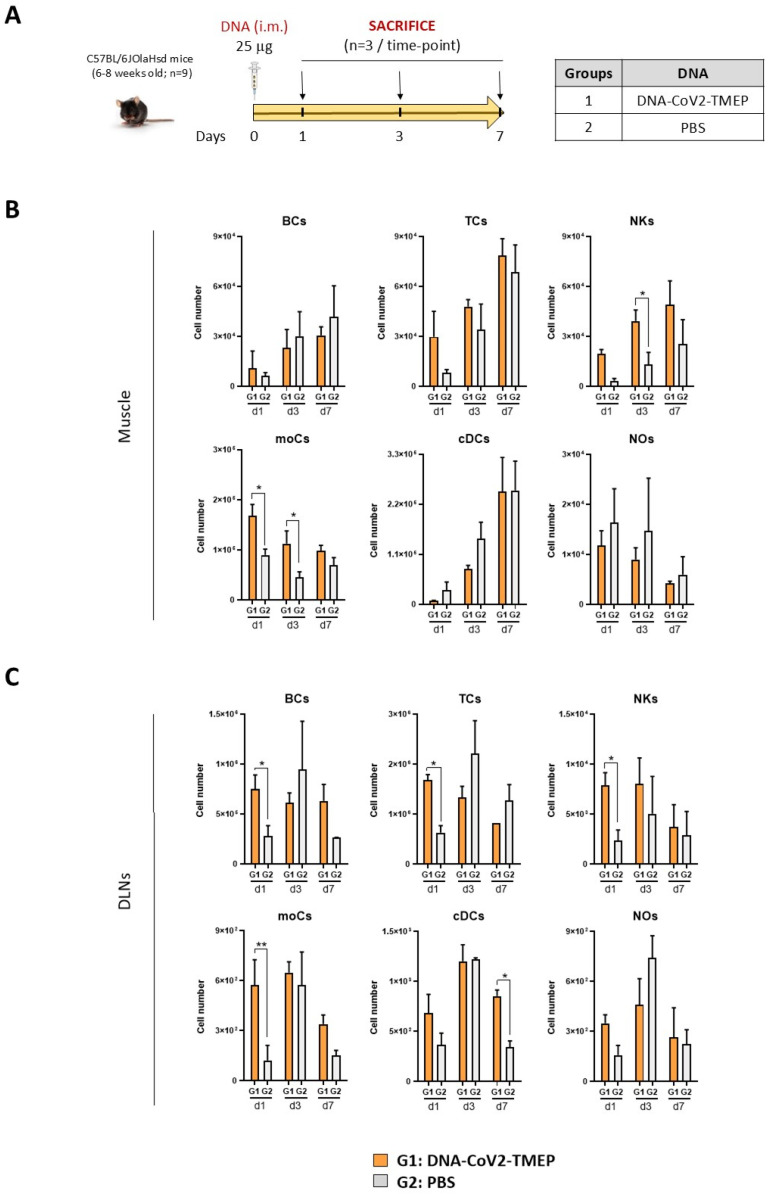
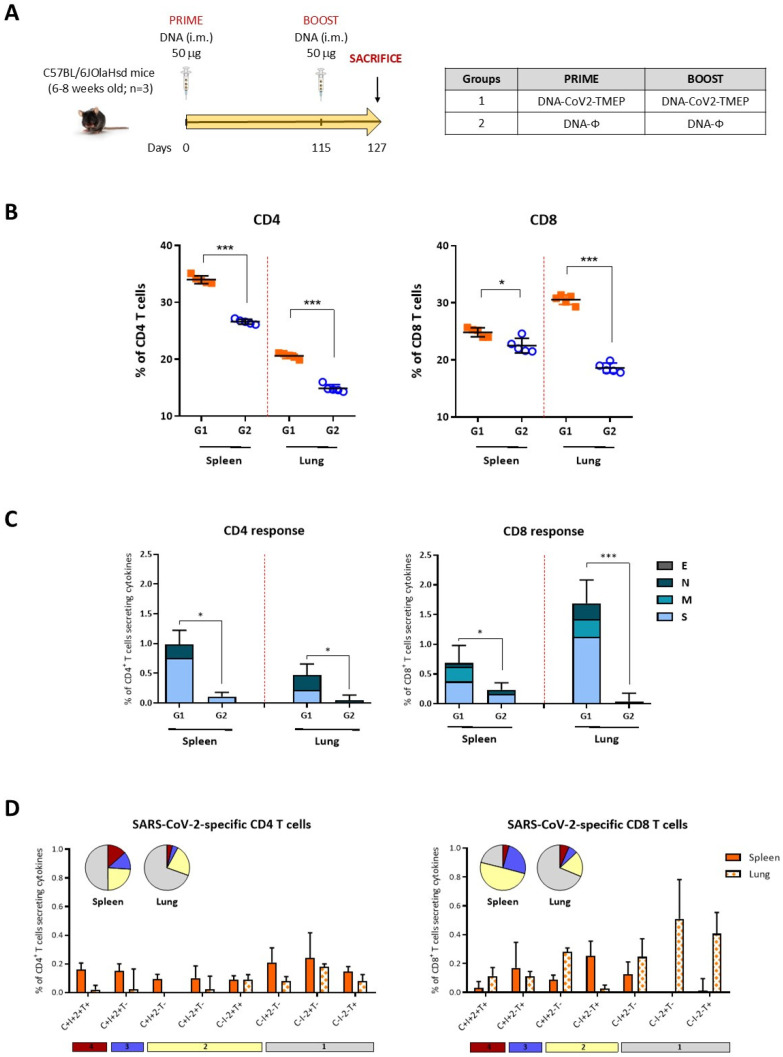
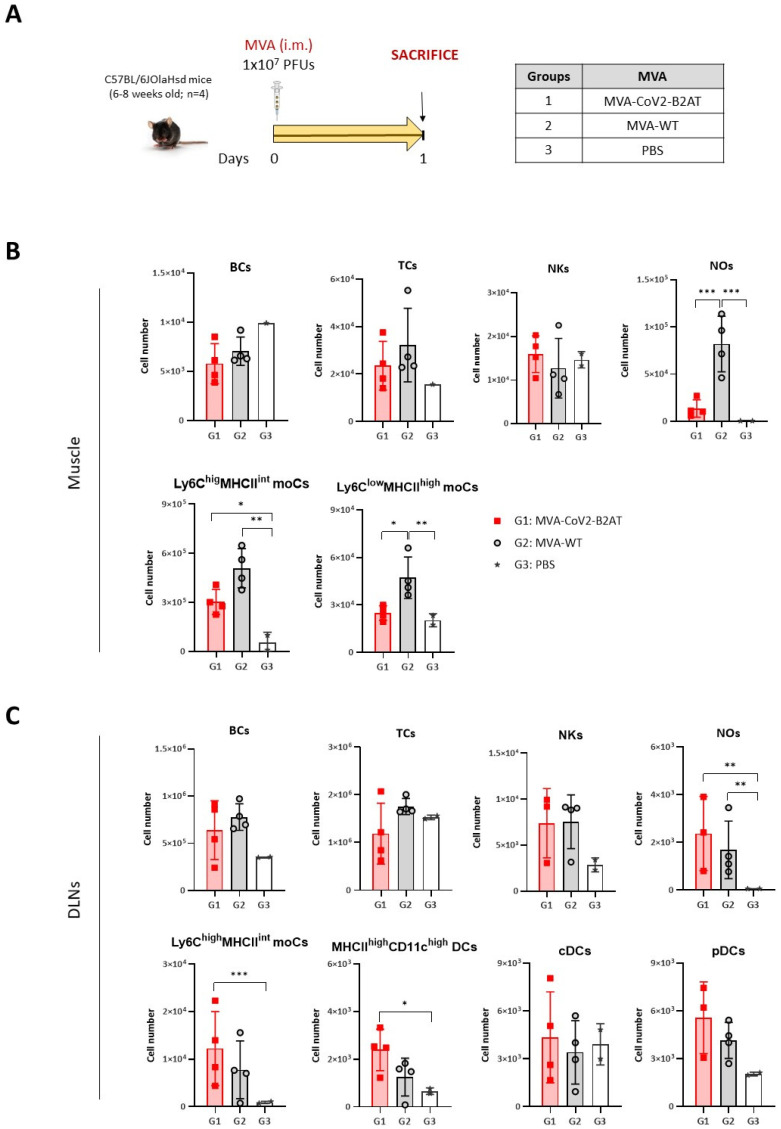
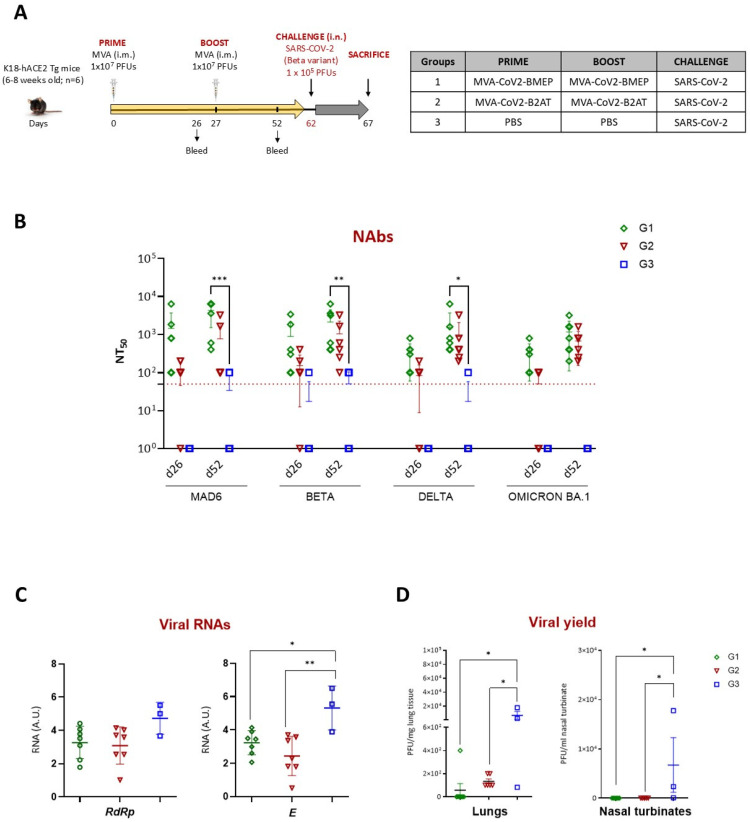
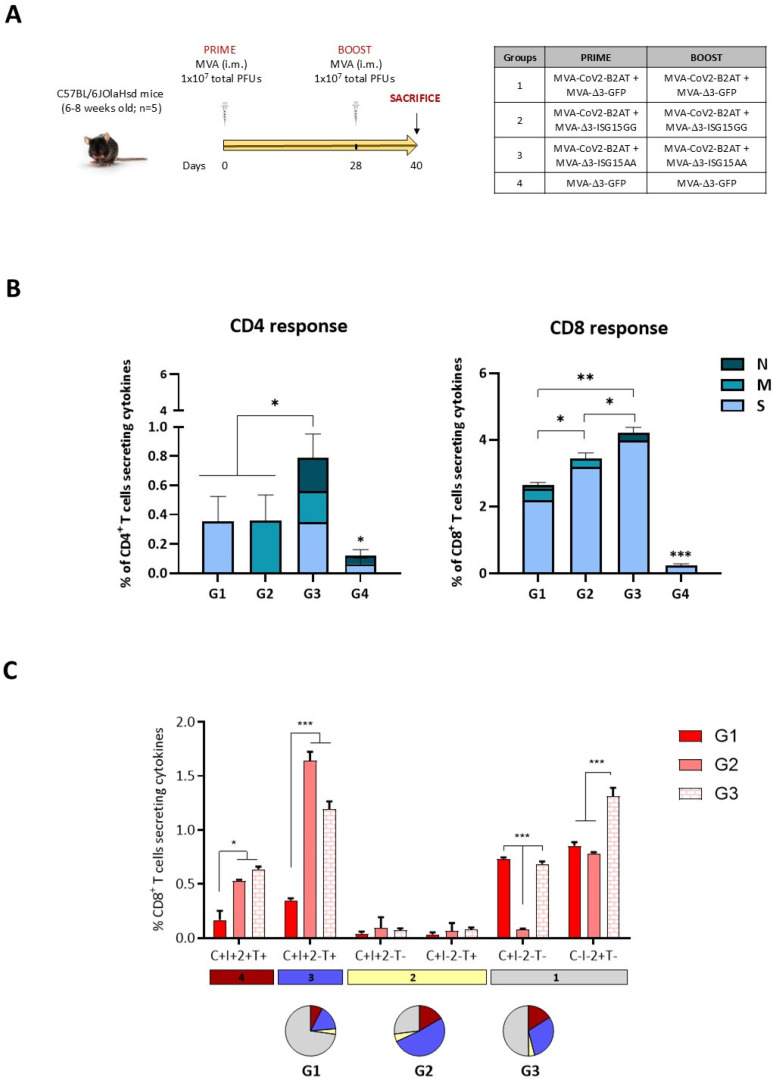
Results: Both candidates were assessed in vitro and in vivo demonstrating their ability to induce robust immune responses. In C57BL/6 mice, DNA-CoV2-TMEP enhanced the recruitment of innate immune cells and stimulated SARS-CoV-2-specific polyfunctional T cells targeting multiple viral proteins. MVA-CoV2-B2AT elicited NAbs against various SARS-CoV-2 variants of concern (VoCs) and reduced viral replication and viral yields against the Beta variant in susceptible K18-hACE2 mice. The combination of MVA-CoV2-B2AT with a mutated ISG15 form as an adjuvant further increased the magnitude, breadth and polyfunctional profile of the response.
Conclusion: These findings underscore the potential of these multiepitopic proteins when expressed from DNA or MVA vectors to provide protection against SARS-CoV-2 and its variants, supporting their further development as next-generation COVID-19 vaccines.
Keywords: DNA and MVA vectors; ISG15 adjuvant; SARS-CoV-2; cellular response; efficacy in mice; immunogenicity; innate response; multiepitopic protein; vaccine.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures
References
-
- Cohen K.W., Linderman S.L., Moodie Z., Czartoski J., Lai L., Mantus G., Norwood C., Nyhoff L.E., Edara V.V., Floyd K., et al. Longitudinal analysis shows durable and broad immune memory after SARS-CoV-2 infection with persisting antibody responses and memory B and T cells. Cell Rep. Med. 2021;2:100354. doi: 10.1016/j.xcrm.2021.100354. - DOI - PMC - PubMed
-
- Meyers L.M., Gutierrez A.H., Boyle C.M., Terry F., McGonnigal B.G., Salazar A., Princiotta M.F., Martin W.D., De Groot A.S., Moise L. Highly conserved, non-human-like, and cross-reactive SARS-CoV-2 T cell epitopes for COVID-19 vaccine design and validation. NPJ Vaccines. 2021;6:71. doi: 10.1038/s41541-021-00331-6. - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
