

Overexpression of LAR1 Suppresses Anthocyanin Biosynthesis by Enhancing Catechin Competition Leading to Promotion of Proanthocyanidin Pathway in Spine Grape (Vitis davidii) Cells
- PMID: 39596158
- PMCID: PMC11594265
- DOI: 10.3390/ijms252212087
Overexpression of LAR1 Suppresses Anthocyanin Biosynthesis by Enhancing Catechin Competition Leading to Promotion of Proanthocyanidin Pathway in Spine Grape (Vitis davidii) Cells
Abstract
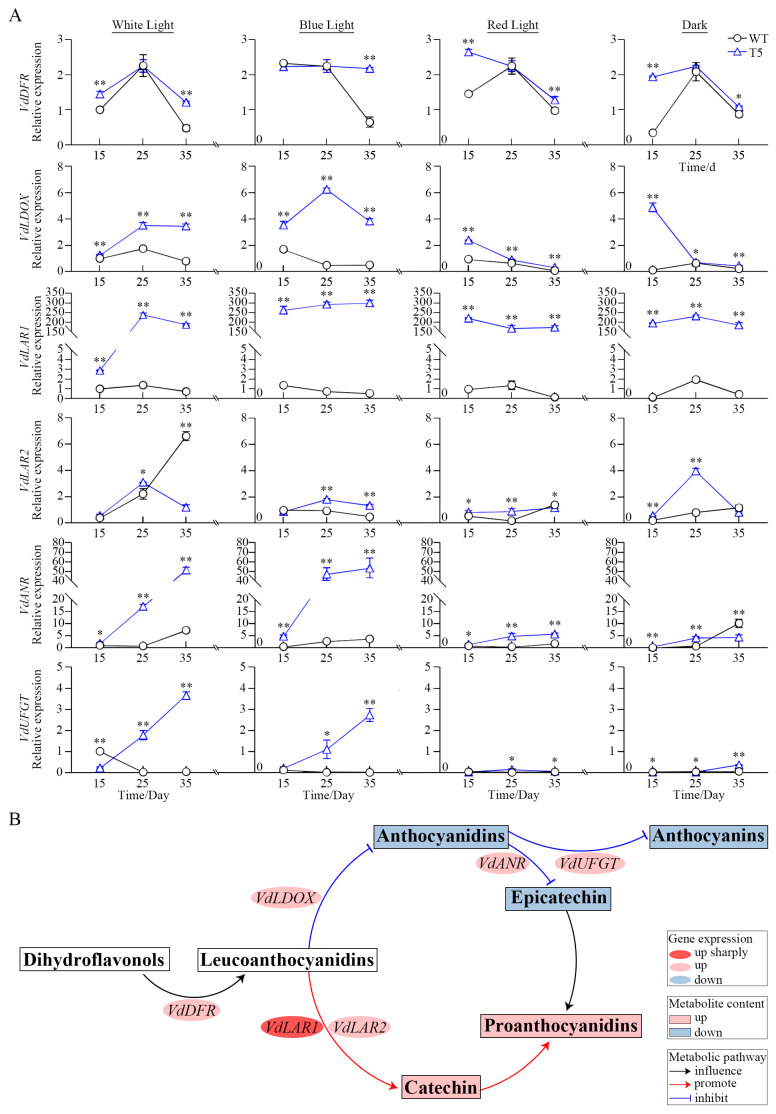
Proanthocyanidins (PAs) are a class of polyphenolic compounds recognized for their potent antioxidant, anti-cancer, anti-inflammatory, and cardioprotective properties. However, the production of PAs from natural sources is often limited by high costs, resource wastage, and environmental damage. In this study, we investigated the overexpression of VdLAR1, along with phenotypic observation, metabolite determination, light quality treatment, and RT-qPCR analysis, in spine grape cells. The results demonstrated a significant increase in the contents of proanthocyanidins and flavonoids in pVdLAR1-overexpressing transgenic cell lines, while anthocyanin levels showed a decreasing trend. Furthermore, the treatment with white and blue light on the T5 cell line resulted in enhanced accumulation of proanthocyanidins, catechins, and flavonoids, whereas anthocyanins and epicatechins exhibited a declining pattern. Thus, short-wavelength light promoted the accumulation of metabolites, with the proanthocyanidin content in the T5 transformed cell line reaching 2512.0 μg/g (FW) during blue light incubation. RT-qPCR analysis revealed that the key genes involved in the biosynthesis of proanthocyanidin and anthocyanin were upregulated in the transgenic spine grape cell lines, with VdLAR1 expression increasing by several hundredfold, far surpassing the expression levels of LDOX and ANR. The VdLAR1 overexpression markedly improved substrate competitiveness within the metabolic pathway, promoting catechin biosynthesis while inhibiting the production of epicatechins and anthocyanins. This finding provides compelling evidence that LAR1 is a crucial gene for catechin biosynthesis. This research establishes both theoretical and practical foundations for the regulation and development of natural proanthocyanidins, addressing issues related to high costs, safety concerns, resource wastage, and environmental damage associated with their production.
Keywords: leucoanthocyanin reductase (LAR); light quality; overexpression; proanthocyanidins; spine grape (Vitis davidii).
Conflict of interest statement
The authors declare no conflicts of interest.
Figures




Similar articles
-
Targeted regulation of 5-aminolevulinic acid enhances flavonoids, anthocyanins and proanthocyanidins accumulation in Vitis davidii callus.BMC Plant Biol. 2024 Oct 10;24(1):944. doi: 10.1186/s12870-024-05667-4. BMC Plant Biol. 2024. PMID: 39385100 Free PMC article.
-
Identification of leucoanthocyanidin reductase and anthocyanidin reductase genes involved in proanthocyanidin biosynthesis in Malus crabapple plants.Plant Physiol Biochem. 2019 Jun;139:141-151. doi: 10.1016/j.plaphy.2019.03.003. Epub 2019 Mar 8. Plant Physiol Biochem. 2019. PMID: 30889479
-
VvLAR1 and VvLAR2 Are Bifunctional Enzymes for Proanthocyanidin Biosynthesis in Grapevine.Plant Physiol. 2019 Jul;180(3):1362-1374. doi: 10.1104/pp.19.00447. Epub 2019 May 15. Plant Physiol. 2019. PMID: 31092697 Free PMC article.
-
Berry skin development in Norton grape: distinct patterns of transcriptional regulation and flavonoid biosynthesis.BMC Plant Biol. 2011 Jan 10;11:7. doi: 10.1186/1471-2229-11-7. BMC Plant Biol. 2011. PMID: 21219654 Free PMC article.
-
The biological functions of proanthocyanidin and its application in pig production.Front Vet Sci. 2025 Mar 12;12:1565501. doi: 10.3389/fvets.2025.1565501. eCollection 2025. Front Vet Sci. 2025. PMID: 40144517 Free PMC article. Review.
Cited by
-
The Strategy and Application of Gene Attenuation in Metabolic Engineering.Microorganisms. 2025 Apr 17;13(4):927. doi: 10.3390/microorganisms13040927. Microorganisms. 2025. PMID: 40284763 Free PMC article. Review.
-
Occurrence, Biosynthesis, and Health Benefits of Anthocyanins in Rice and Barley.Int J Mol Sci. 2025 Jun 27;26(13):6225. doi: 10.3390/ijms26136225. Int J Mol Sci. 2025. PMID: 40649998 Free PMC article. Review.
References
-
- Ramsay A., Mueller-Harvey I. Procyanidins from Averrhoa bilimbi fruits and leaves. J. Food Compos. Anal. 2016;47:16–20. doi: 10.1016/j.jfca.2015.12.004. - DOI
-
- Qi Q., Chu M., Yu X., Xie Y., Li Y., Du Y., Liu X., Zhang Z., Shi J., Yan N. Anthocyanins and Proanthocyanidins: Chemical Structures, Food Sources, Bioactivities, and Product Development. Food Rev. Int. 2022;39:4581–4609. doi: 10.1080/87559129.2022.2029479. - DOI
-
- Wu Q., Li S.-Y., Yang T., Xiao J., Chu Q.-M., Li T., Xie B.-J., Sun Z.-D. Inhibitory effect of lotus seedpod oligomeric procyanidins on advanced glycation end product formation in a lactose–lysine model system. Electron. J. Biotechnol. 2015;18:68–76. doi: 10.1016/j.ejbt.2014.10.005. - DOI
MeSH terms
Substances
Grants and funding
- 2022J01482/Fujian provincial natural science foundation
- GJYS202401/agricultural science and technology special project of Fujian Academy of Agricultural Sciences
- Minkexiexue 2023-13/Fujian Province technology and economic integration service platform of characteristic agricultural product processing
LinkOut - more resources
Full Text Sources
Miscellaneous
