

Mutational Analysis of Substrate Recognition in Trypsin-like Protease Cocoonase: Protein Memory Induced by Alterations in Substrate-Binding Site
- PMID: 39598863
- PMCID: PMC11597625
- DOI: 10.3390/molecules29225476
Mutational Analysis of Substrate Recognition in Trypsin-like Protease Cocoonase: Protein Memory Induced by Alterations in Substrate-Binding Site
Abstract
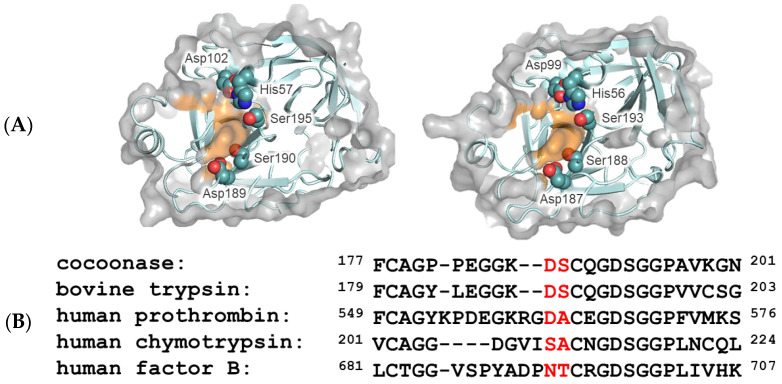
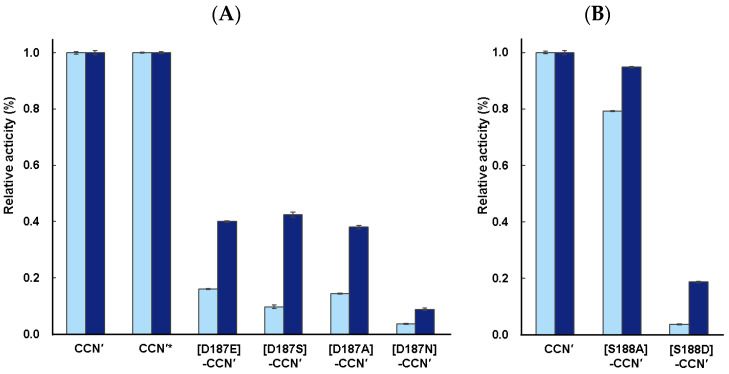
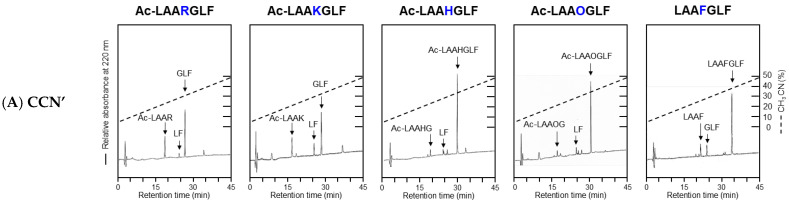
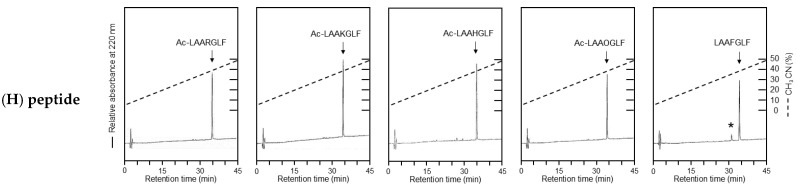
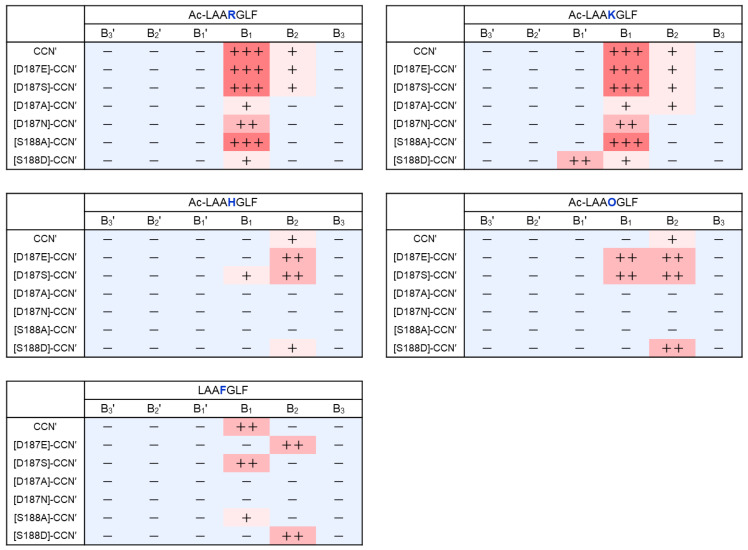
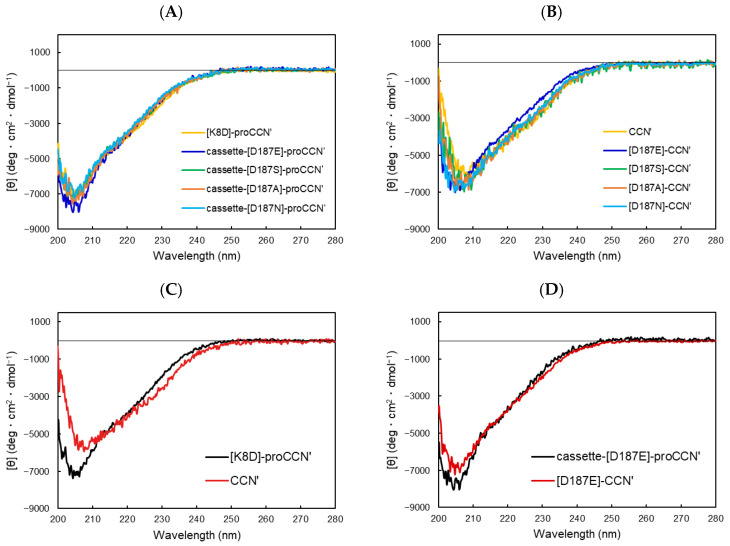
To investigate the substrate recognition mechanism of trypsin-like protease cocoonase (CCN), mutational analyses were conducted at key substrate recognition sites, Asp187 and Ser188, and their effects on substrate specificity and enzymatic activity were evaluated. Mutants with the Asp187 substitution exhibited a significant reduction in catalytic activity compared with the wild-type enzyme, whereas the Ser188 mutants displayed a comparatively minor effect on activity. This indicates that Asp187 plays a crucial role in catalytic function, whereas Ser188 serves a complementary role in substrate recognition. Interestingly, the substitution of the Asp187 to Glu or Ser caused novel substrate specificities, resulting in the recognition of Orn and His residues. In addition, when Asp187 and Ser188 were substituted with acidic residues (Glu or Asp), both the precursor proCCN and mature CCN proteins retained highly similar secondary and tertiary structures. This reveals that the structural characteristics of precursor proteins are maintained in the mature proteins, potentially influencing substrate recognition and catalytic function. These findings suggest that the pro-regions of these mutants interact much more tightly with the mature enzyme than in the wild-type CCN. These results provide fruitful insights into the structural determinants governing substrate recognition in enzyme variants.
Keywords: chaperone; chymotrypsin; cocoonase; folding; substrate binding.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures


References
-
- Huber R., Bode W. Structural basis of the activation and action of trypsin. Acc. Chem. Res. 1978;11:114–122. doi: 10.1021/ar50123a006. - DOI
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
