

Seeding-competent TDP-43 persists in human patient and mouse muscle
- PMID: 39602508
- PMCID: PMC11812673
- DOI: 10.1126/scitranslmed.adp5730
Seeding-competent TDP-43 persists in human patient and mouse muscle
Abstract
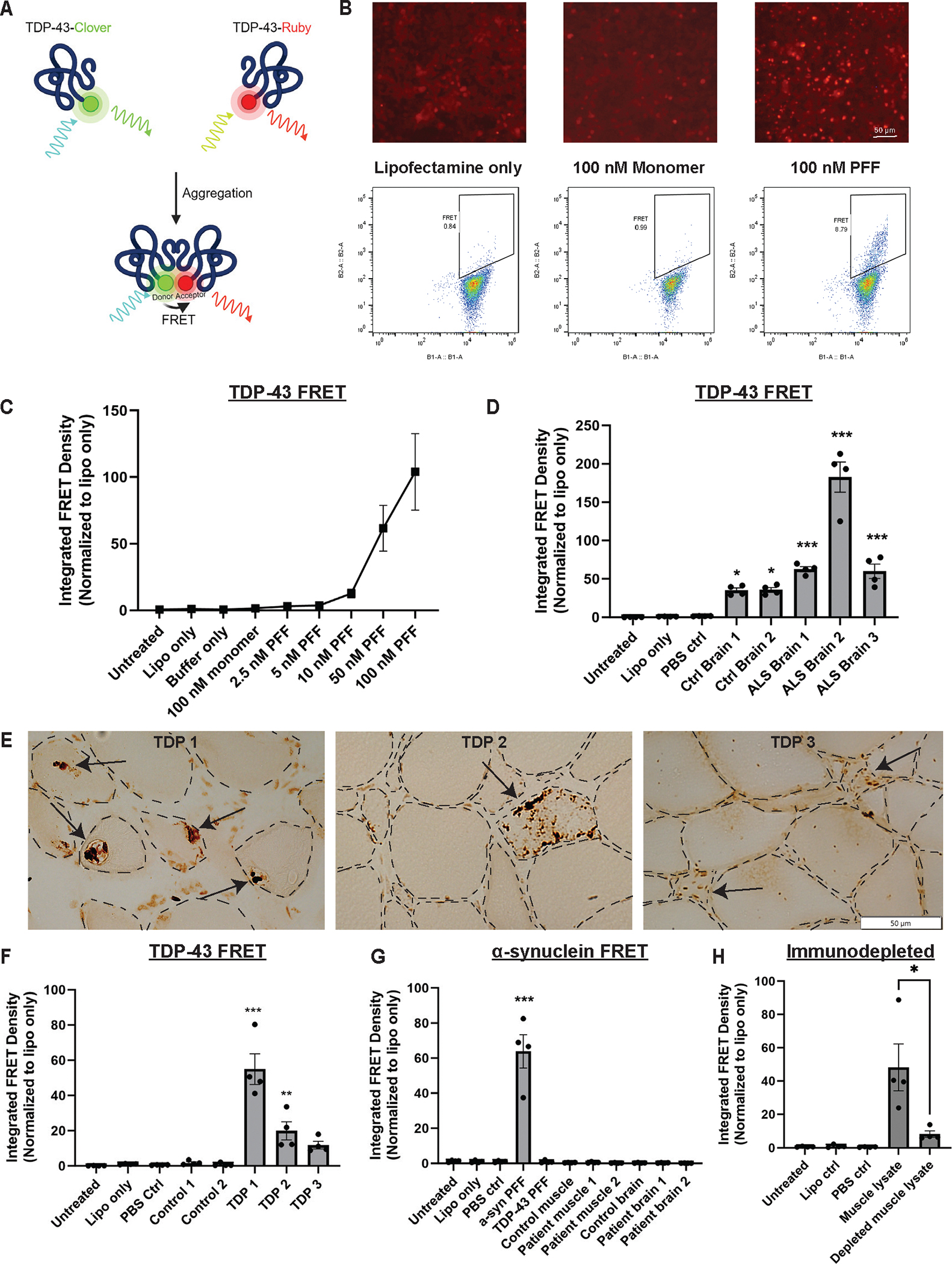
TAR DNA binding protein 43 (TDP-43) is an RNA binding protein that accumulates as aggregates in the central nervous systems of some patients with neurodegenerative diseases. However, TDP-43 aggregation is also a sensitive and specific pathologic feature found in a family of degenerative muscle diseases termed inclusion body myopathy. TDP-43 aggregates from amyotrophic lateral sclerosis (ALS) and frontotemporal dementia brain lysates may serve as self-templating aggregate seeds in vitro and in vivo, supporting a prion-like spread from cell to cell. Whether a similar process occurs in patient muscle is not clear. We developed a mouse model of inducible, muscle-specific cytoplasmic localized TDP-43. These mice develop muscle weakness with robust accumulation of insoluble and phosphorylated sarcoplasmic TDP-43, leading to eosinophilic inclusions, altered proteostasis, and changes in TDP-43-related RNA processing that resolve with the removal of doxycycline. Skeletal muscle lysates from these mice also have seeding-competent TDP-43, as determined by a FRET-based biosensor, that persists for weeks upon resolution of TDP-43 aggregate pathology. Human muscle biopsies with TDP-43 pathology also contain TDP-43 aggregate seeds. Using lysates from muscle biopsies of patients with sporadic inclusion body myositis (IBM), immune-mediated necrotizing myopathy (IMNM), and ALS, we found that TDP-43 seeding capacity was specific to IBM. TDP-43 seeding capacity anticorrelated with TDP-43 aggregate and vacuole abundance. These data support that TDP-43 aggregate seeds are present in IBM skeletal muscle and represent a unique TDP-43 pathogenic species not previously appreciated in human muscle disease.
Conflict of interest statement
Competing interests:
None to disclose.
Figures







Update of
-
Seeding competent TDP-43 persists in human patient and mouse muscle.bioRxiv [Preprint]. 2024 Apr 4:2024.04.03.587918. doi: 10.1101/2024.04.03.587918. bioRxiv. 2024. Update in: Sci Transl Med. 2024 Nov 27;16(775):eadp5730. doi: 10.1126/scitranslmed.adp5730. PMID: 38617354 Free PMC article. Updated. Preprint.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
