

Insights into RNA-mediated pathology in new mouse models of Huntington's disease
- PMID: 39604147
- PMCID: PMC11602643
- DOI: 10.1096/fj.202401465R
Insights into RNA-mediated pathology in new mouse models of Huntington's disease
Abstract
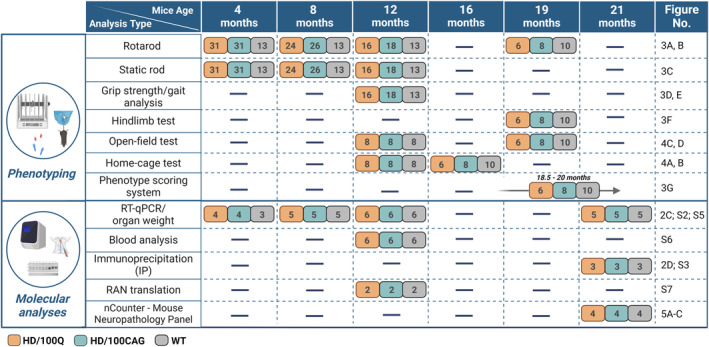
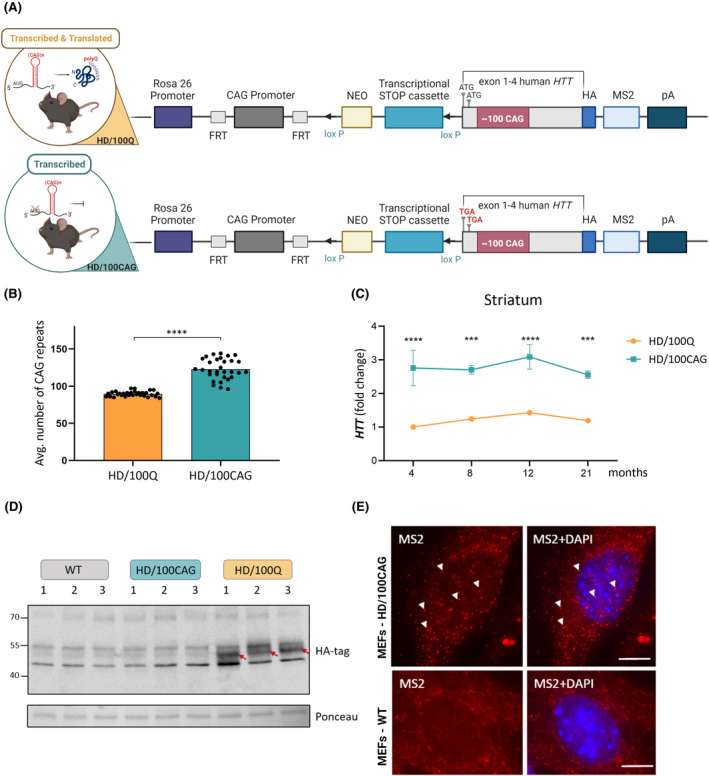
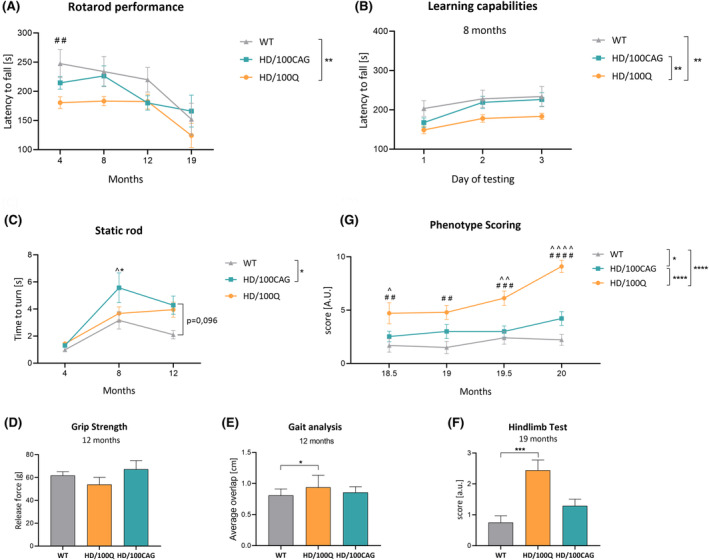
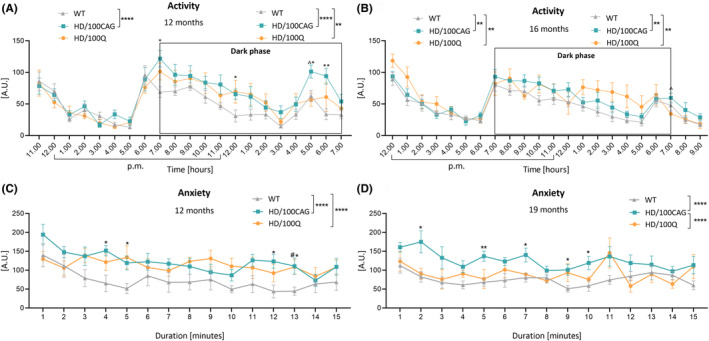
Huntington's disease (HD) is a neurodegenerative polyglutamine (polyQ) disease resulting from the expansion of CAG repeats located in the ORF of the huntingtin gene (HTT). The extent to which mutant mRNA-driven disruptions contribute to HD pathogenesis, particularly in comparison to the dominant mechanisms related to the gain-of-function effects of the mutant polyQ protein, is still debatable. To evaluate this contribution in vivo, we generated two mouse models through a knock-in strategy at the Rosa26 locus. These models expressed distinct variants of human mutant HTT cDNA fragment: a translated variant (HD/100Q model, serving as a reference) and a nontranslated variant (HD/100CAG model). The cohorts of animals were subjected to a broad spectrum of molecular, behavioral, and cognitive analysis for 21 months. Behavioral testing revealed alterations in both models, with the HD/100Q model exhibiting late disease phenotype. The rotarod, static rod, and open-field tests showed some motor deficits in HD/100CAG and HD/100Q model mice during the light phase, while ActiMot indicated hyperkinesis during the dark phase. Both models also exhibited certain gene deregulations in the striatum that are related to disrupted pathways and phenotype alterations observed in HD. In conclusion, we provide in vivo evidence for a minor contributory role of mutant RNA in HD pathogenesis. The separated effects resulting from the presence of mutant RNA in the HD/100CAG model led to less severe but, to some extent, similar types of impairments as in the HD/100Q model. Increased anxiety was one of the most substantial effects caused by mutant HTT RNA.
Keywords: Huntington's disease; RNA toxicity; mouse model; neurodegenerative diseases; polyglutamine disorders.
© 2024 The Author(s). The FASEB Journal published by Wiley Periodicals LLC on behalf of Federation of American Societies for Experimental Biology.
Figures
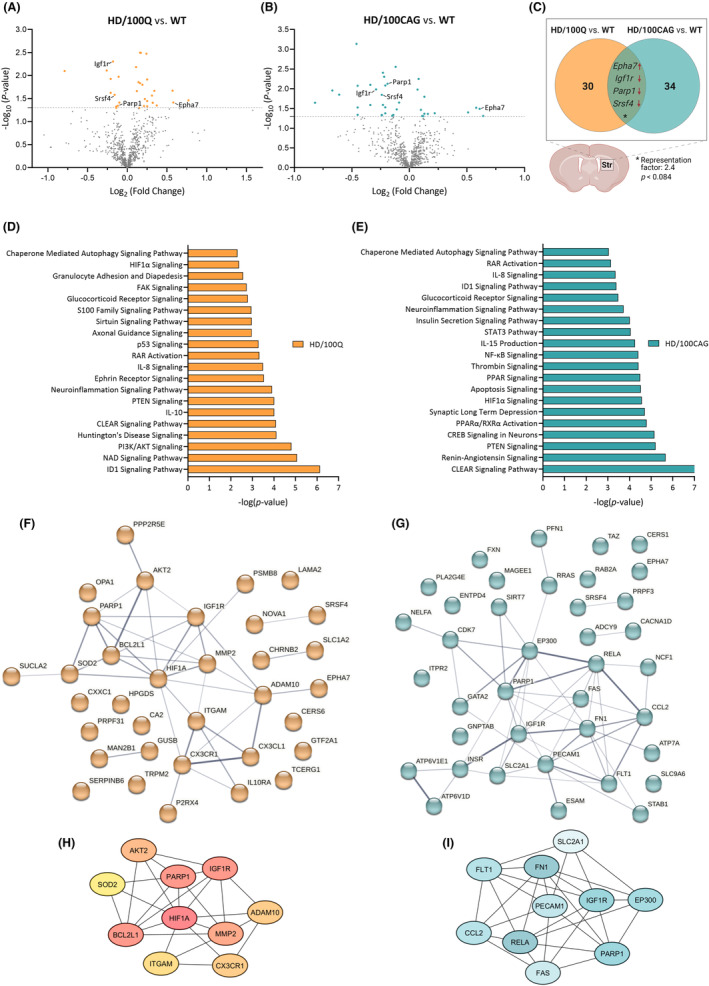
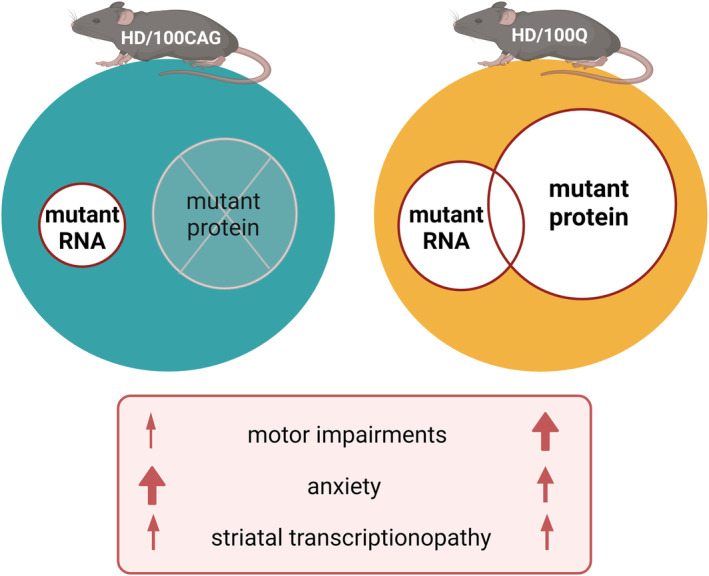
References
-
- Ghosh R, Tabrizi SJ. Huntington disease. Handb Clin Neurol. 2018;147:255‐278. - PubMed
-
- MacDonald ME, Ambrose CM, Duyao MP, et al. A novel gene containing a trinucleotide repeat that is expanded and unstable on Huntington's disease chromosomes. The Huntington's disease collaborative research group. Cell. 1993;72:971‐983. - PubMed
-
- Tabrizi SJ, Flower MD, Ross CA, Wild EJ. Huntington disease: new insights into molecular pathogenesis and therapeutic opportunities. Nat Rev Neurol. 2020;16:529‐546. - PubMed
-
- Fusilli C, Migliore S, Mazza T, et al. Biological and clinical manifestations of juvenile Huntington's disease: a retrospective analysis. Lancet Neurol. 2018;17:986‐993. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
