

Fibrous corona is reduced in cancer cell lines that attenuate microtubule nucleation from kinetochores
- PMID: 39604214
- PMCID: PMC11786318
- DOI: 10.1111/cas.16406
Fibrous corona is reduced in cancer cell lines that attenuate microtubule nucleation from kinetochores
Abstract
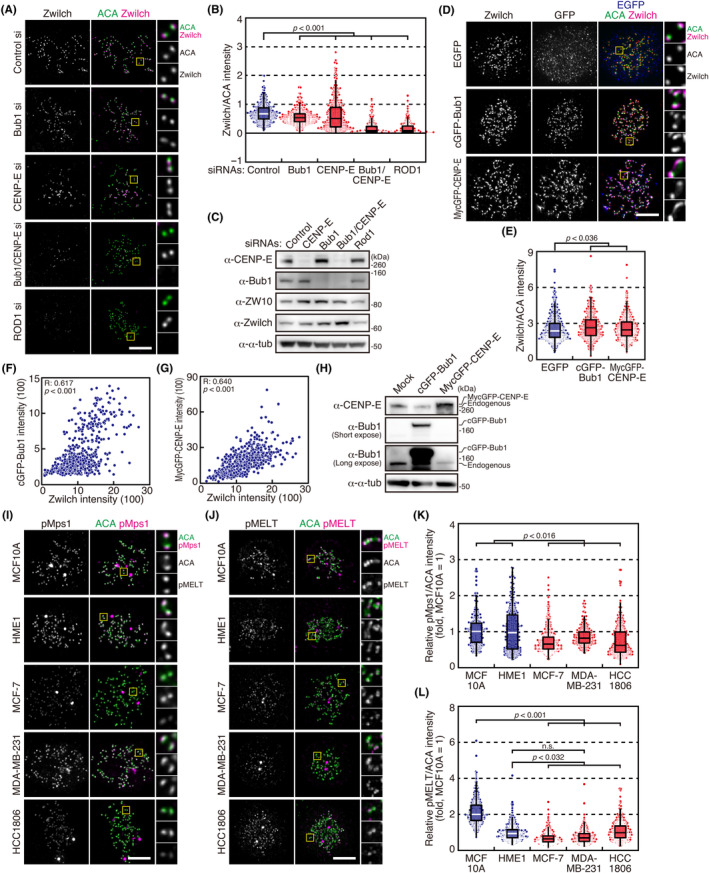
Most cancer cells show increased chromosome missegregation, known as chromosomal instability (CIN), which promotes cancer progression and drug resistance. The underlying causes of CIN in cancer cells are not fully understood. Here we found that breast cancer cell lines show a reduced kinetochore localization of ROD, ZW10, and Zwilch, components of the fibrous corona, compared with non-transformed breast epithelial cell lines. The fibrous corona is a structure formed on kinetochores before their end-on attachment to microtubules and plays a role in efficient kinetochore capture and the spindle assembly checkpoint. The reduction in the fibrous corona was not due to reduced expression levels of the fibrous corona components or to a reduction in outer kinetochore components. Kinetochore localization of Bub1 and CENP-E, which play a role in the recruitment of the fibrous corona to kinetochores, was reduced in cancer cell lines, presumably due to reduced activity of Mps1, which is required for their recruitment to kinetochores through phosphorylating KNL1. Increasing kinetochore localization of Bub1 and CENP-E in cancer cells restored the level of the fibrous corona. Cancer cell lines showed a reduced capacity to nucleate microtubules from kinetochores, which was recently shown to be dependent on the fibrous corona, and increasing kinetochore localization of Bub1 and CENP-E restored the microtubule nucleation capacity on kinetochores. Our study revealed a distinct feature of cancer cell lines that may be related to CIN.
Keywords: Bub1; CENP‐E; RZZ complex; breast cancer; chromosomal instability; fibrous corona.
© 2024 The Author(s). Cancer Science published by John Wiley & Sons Australia, Ltd on behalf of Japanese Cancer Association.
Conflict of interest statement
The authors have no competing financial interests. Kozo Tanaka is a current Editorial Board Member of
Figures




References
-
- Weaver BA, Cleveland DW. Does aneuploidy cause cancer? Curr Opin Cell Biol. 2006;18:658‐667. - PubMed
-
- Ben‐David U, Amon A. Context is everything: aneuploidy in cancer. Nat Rev Genet. 2020;21:44‐62. - PubMed
-
- Tanaka K, Hirota T. Chromosomal instability: a common feature and a therapeutic target of cancer. Biochim Biophys Acta. 2016;1866:64‐75. - PubMed
-
- Gordon DJ, Resio B, Pellman D. Causes and consequences of aneuploidy in cancer. Nat Rev Genet. 2012;13:189‐203. - PubMed
MeSH terms
Substances
Grants and funding
- 18H04896/Ministry of Education, Culture, Sports, Science and Technology
- 21H05738/Ministry of Education, Culture, Sports, Science and Technology
- 23H04272/Ministry of Education, Culture, Sports, Science and Technology
- Yamaguchi Educational and Scholarship Foundation
- Mochida Memorial Foundation for Medical and Pharmaceutical Research
- JPMJAX2112/Japan Science and Technology Agency
- Takeda Science Foundation
- The Pharmacological Research Foundation. Tokyo
- 15H04368/Japan Society for the Promotion of Science
- 16H06635/Japan Society for the Promotion of Science
- 16K14604/Japan Society for the Promotion of Science
- 18H02434/Japan Society for the Promotion of Science
- 18K15234/Japan Society for the Promotion of Science
- 22H02614/Japan Society for the Promotion of Science
- 23K05629/Japan Society for the Promotion of Science
LinkOut - more resources
Full Text Sources
Medical
Research Materials
