

Engineered transcription-associated Cas9 targeting in eukaryotic cells
- PMID: 39604381
- PMCID: PMC11603292
- DOI: 10.1038/s41467-024-54629-9
Engineered transcription-associated Cas9 targeting in eukaryotic cells
Abstract
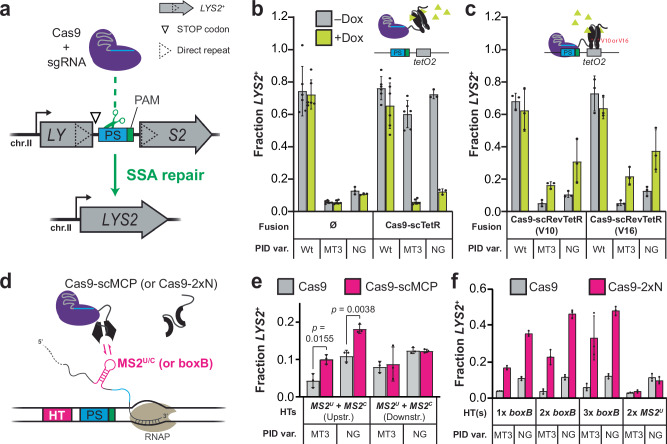
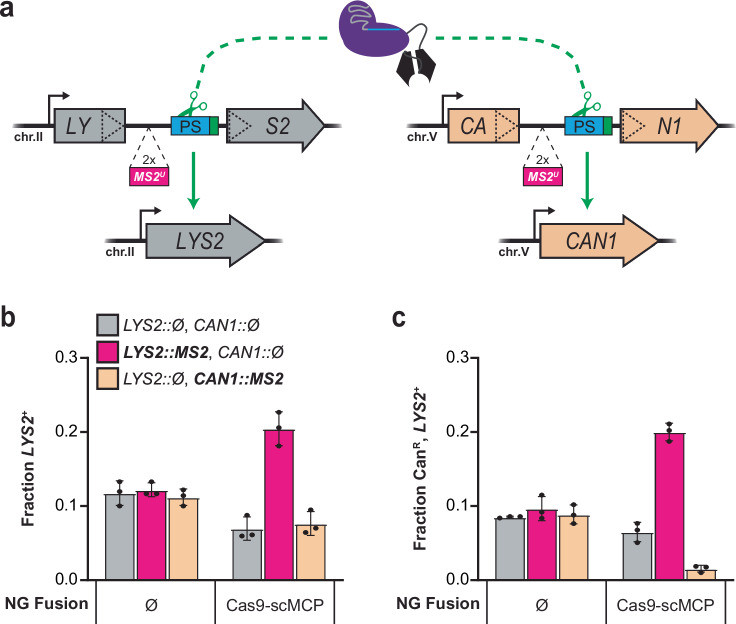
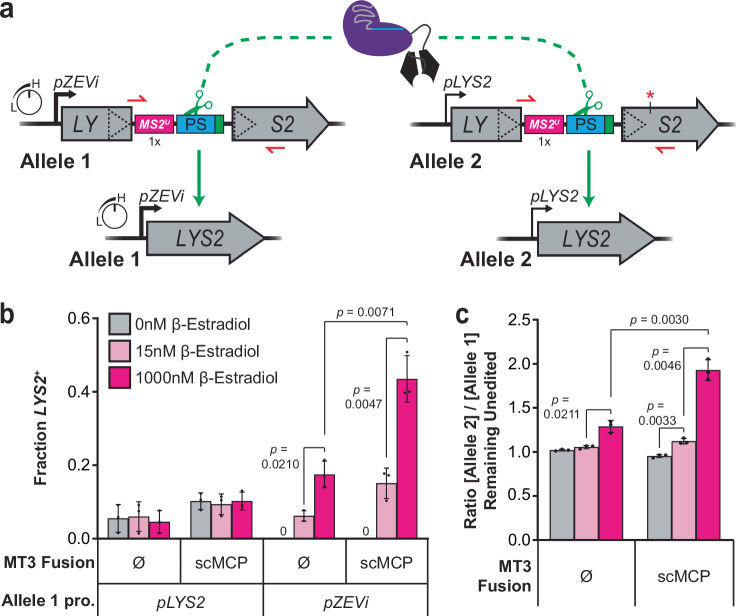
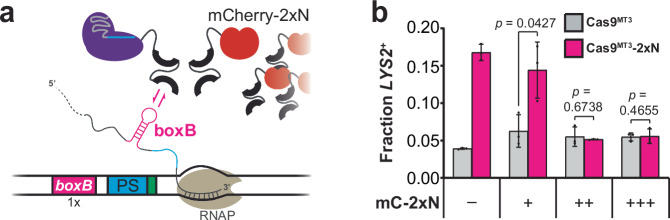
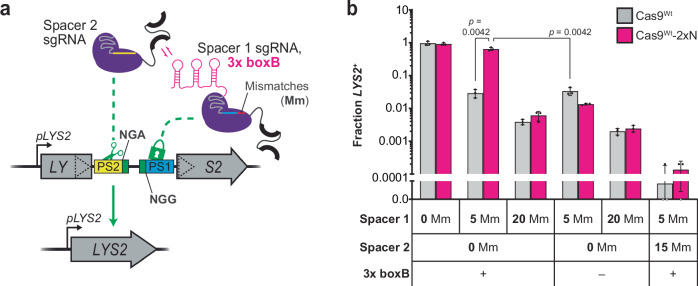
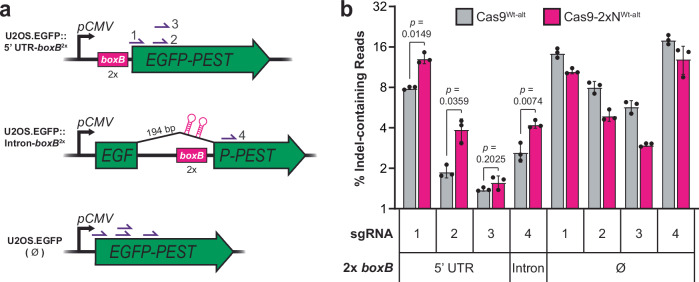
DNA targeting Class 2 CRISPR-Cas effector nucleases, including the well-studied Cas9 proteins, evolved protospacer-adjacent motif (PAM) and guide RNA interactions that sequentially license their binding and cleavage activities at protospacer target sites. Both interactions are nucleic acid sequence specific but function constitutively; thus, they provide intrinsic spatial control over DNA targeting activities but naturally lack temporal control. Here we show that engineered Cas9 fusion proteins which bind to nascent RNAs near a protospacer can facilitate spatiotemporal coupling between transcription and DNA targeting at that protospacer: Transcription-associated Cas9 Targeting (TraCT). Engineered TraCT is enabled in eukaryotic yeast or human cells when suboptimal PAM interactions limit basal activity and when one or more nascent RNA substrates are still tethered to the actively transcribed target DNA in cis. Using yeast, we further show that this phenomenon can be applied for selective editing at one of two identical targets in distinct gene loci, or, in diploid allelic loci that are differentially transcribed. Our work demonstrates that temporal control over Cas9's targeting activity at specific DNA sites may be engineered without modifying Cas9's core domains and guide RNA components or their expression levels. More broadly, it establishes co-transcriptional RNA binding as a cis-acting mechanism that can conditionally stimulate CRISPR-Cas DNA targeting in eukaryotic cells.
© 2024. The Author(s).
Conflict of interest statement
Competing interests: Jef Boeke is a Founder and Director of CDI Labs, Inc., a Founder of and consultant to Opentrons LabWorks/Neochromosome, Inc, and serves or served on the Scientific Advisory Board of the following: CZ Biohub New York, LLC, Logomix, Inc., Modern Meadow, Inc., Rome Therapeutics, Inc., SeaHub, Seattle, WA, Tessera Therapeutics, Inc. and the Wyss Institute. Marcus Noyes is a founder of TBG Therapeutics, Inc. New York University filed a provisional patent application (No. 63/582,731) for findings described in this work, with Gregory Goldberg, Marcus Noyes, and Jef Boeke listed as inventors. The other authors declare no competing interests.
Figures

Update of
-
Engineered transcription-associated Cas9 targeting in eukaryotic cells.bioRxiv [Preprint]. 2024 Aug 18:2023.09.18.558319. doi: 10.1101/2023.09.18.558319. bioRxiv. 2024. Update in: Nat Commun. 2024 Nov 27;15(1):10287. doi: 10.1038/s41467-024-54629-9. PMID: 37781609 Free PMC article. Updated. Preprint.
References
Publication types
MeSH terms
Substances
Associated data
Grants and funding
- F32GM137482/U.S. Department of Health & Human Services | NIH | National Institute of General Medical Sciences (NIGMS)
- R01 GM133936/GM/NIGMS NIH HHS/United States
- RM1HG009491/U.S. Department of Health & Human Services | NIH | National Human Genome Research Institute (NHGRI)
- RM1 HG009491/HG/NHGRI NIH HHS/United States
- R01 GM118851/GM/NIGMS NIH HHS/United States
- F32 GM137482/GM/NIGMS NIH HHS/United States
- K99 GM147604/GM/NIGMS NIH HHS/United States
- R01GM118851/U.S. Department of Health & Human Services | NIH | National Institute of General Medical Sciences (NIGMS)
- K99GM147604/U.S. Department of Health & Human Services | NIH | National Institute of General Medical Sciences (NIGMS)
LinkOut - more resources
Full Text Sources
Miscellaneous
