

Remodelling of Rea1 linker domain drives the removal of assembly factors from pre-ribosomal particles
- PMID: 39604383
- PMCID: PMC11603028
- DOI: 10.1038/s41467-024-54698-w
Remodelling of Rea1 linker domain drives the removal of assembly factors from pre-ribosomal particles
Abstract
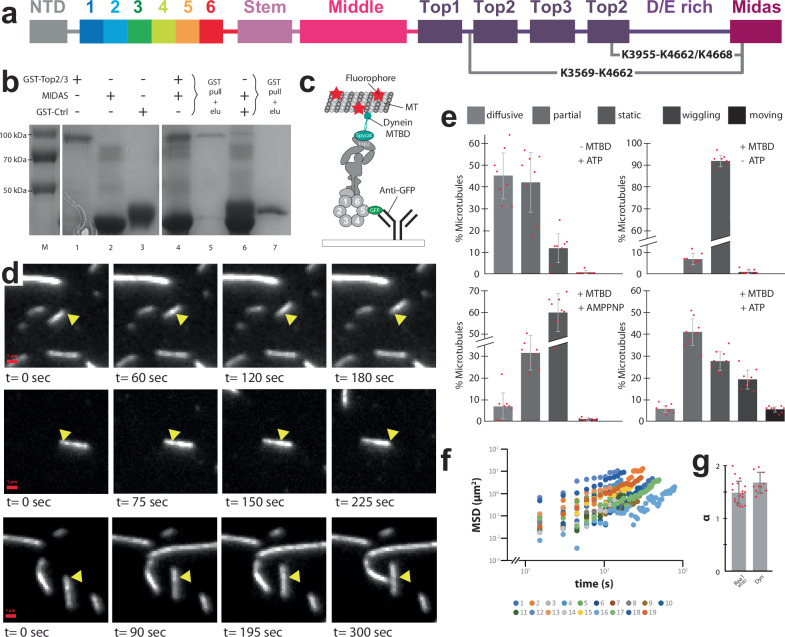
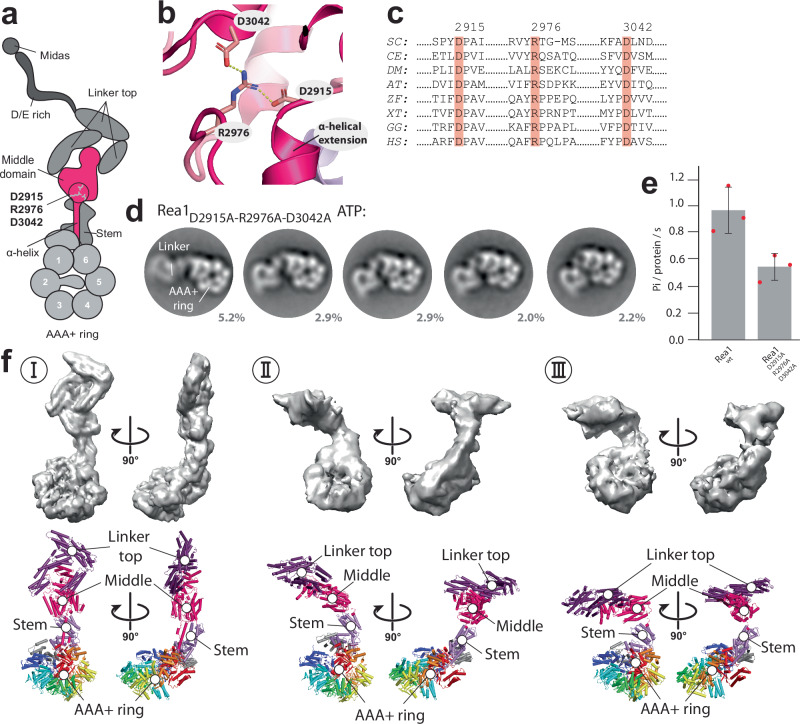
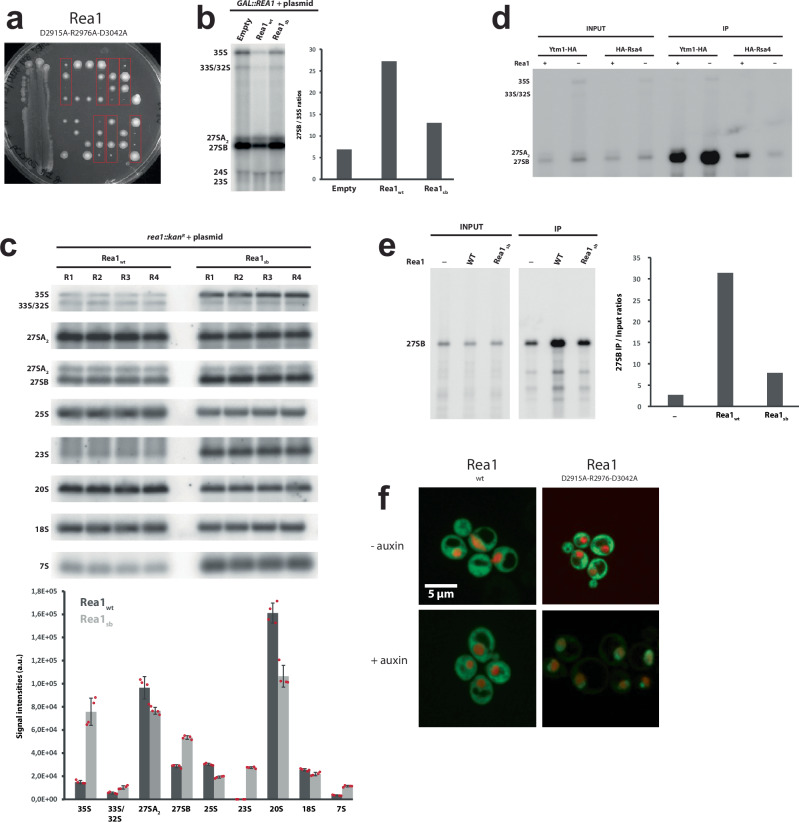
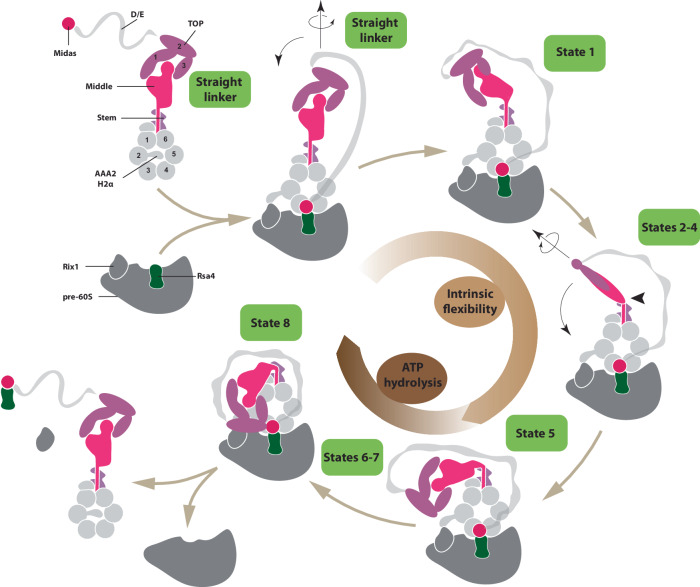
The ribosome maturation factor Rea1 (or Midasin) catalyses the removal of assembly factors from large ribosomal subunit precursors and promotes their export from the nucleus to the cytosol. Rea1 consists of nearly 5000 amino-acid residues and belongs to the AAA+ protein family. It consists of a ring of six AAA+ domains from which the ≈1700 amino-acid residue linker emerges that is subdivided into stem, middle and top domains. A flexible and unstructured D/E rich region connects the linker top to a MIDAS (metal ion dependent adhesion site) domain, which is able to bind the assembly factor substrates. Despite its key importance for ribosome maturation, the mechanism driving assembly factor removal by Rea1 is still poorly understood. Here we demonstrate that the Rea1 linker is essential for assembly factor removal. It rotates and swings towards the AAA+ ring following a complex remodelling scheme involving nucleotide independent as well as nucleotide dependent steps. ATP-hydrolysis is required to engage the linker with the AAA+ ring and ultimately with the AAA+ ring docked MIDAS domain. The interaction between the linker top and the MIDAS domain allows direct force transmission for assembly factor removal.
© 2024. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures
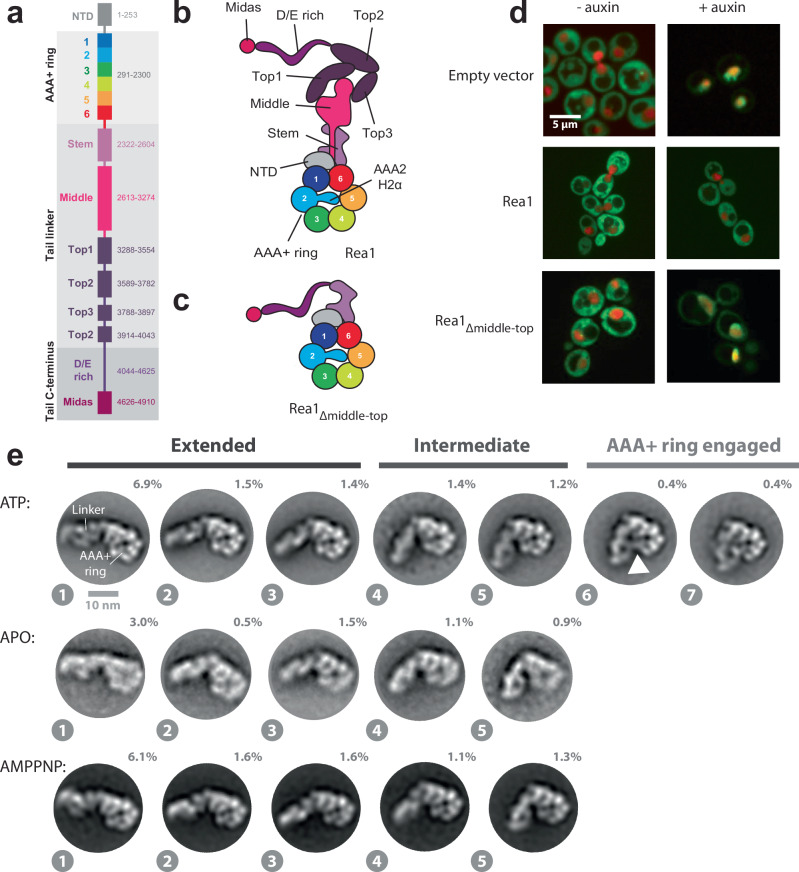
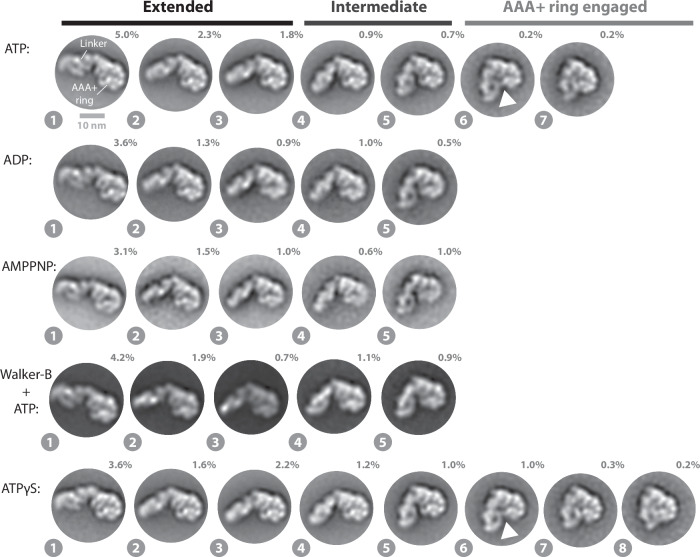
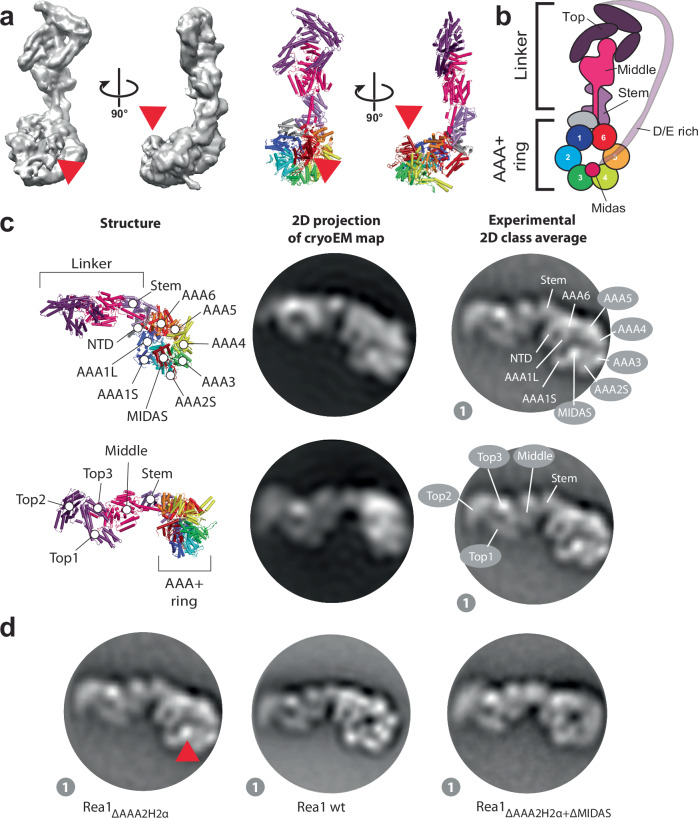
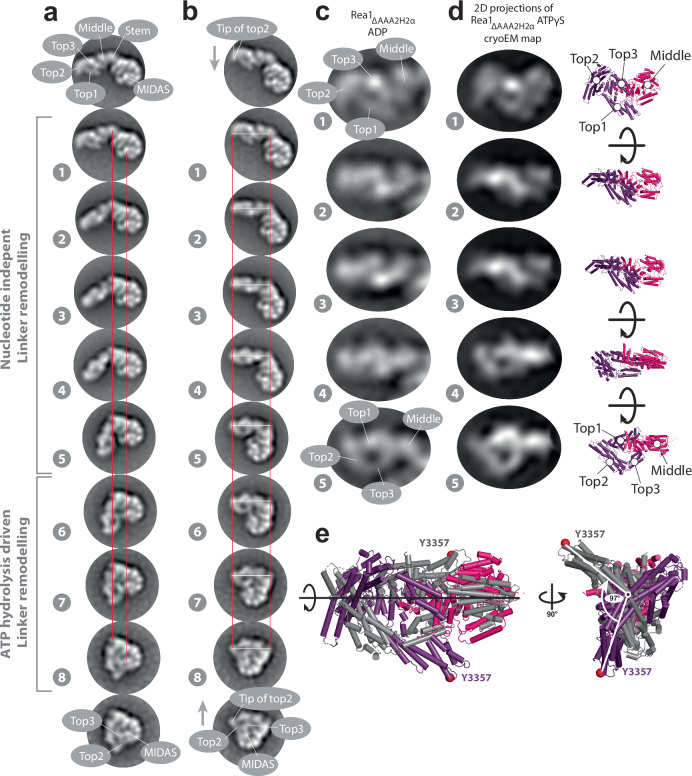
References
-
- Thomson, E., Ferreira-Cerca, S. & Hurt, E. Eukaryotic ribosome biogenesis at a glance. J. Cell Sci.126, 4815–4821 (2013). - PubMed
-
- Kressler, D., Hurt, E. & Bassler, J. Driving ribosome assembly. Biochim Biophys. Acta1803, 673–683 (2010). - PubMed
-
- Harnpicharnchai, P. et al. Composition and functional characterization of yeast 66S ribosome assembly intermediates. Mol. Cell8, 505–515 (2001). - PubMed
-
- Venema, J. & Tollervey, D. Ribosome synthesis in Saccharomyces cerevisiae. Annu Rev. Genet33, 261–311 (1999). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
