

This is a preprint.
Single-cell analyses reveal that monocyte gene expression profiles influence HIV-1 reservoir size in acutely treated cohorts
- PMID: 39605411
- PMCID: PMC11601329
- DOI: 10.1101/2024.11.12.623270
Single-cell analyses reveal that monocyte gene expression profiles influence HIV-1 reservoir size in acutely treated cohorts
Update in
-
Single-cell analyses identify monocyte gene expression profiles that influence HIV-1 reservoir size in acutely treated cohorts.Nat Commun. 2025 May 29;16(1):4975. doi: 10.1038/s41467-025-59833-9. Nat Commun. 2025. PMID: 40442100 Free PMC article.
Abstract
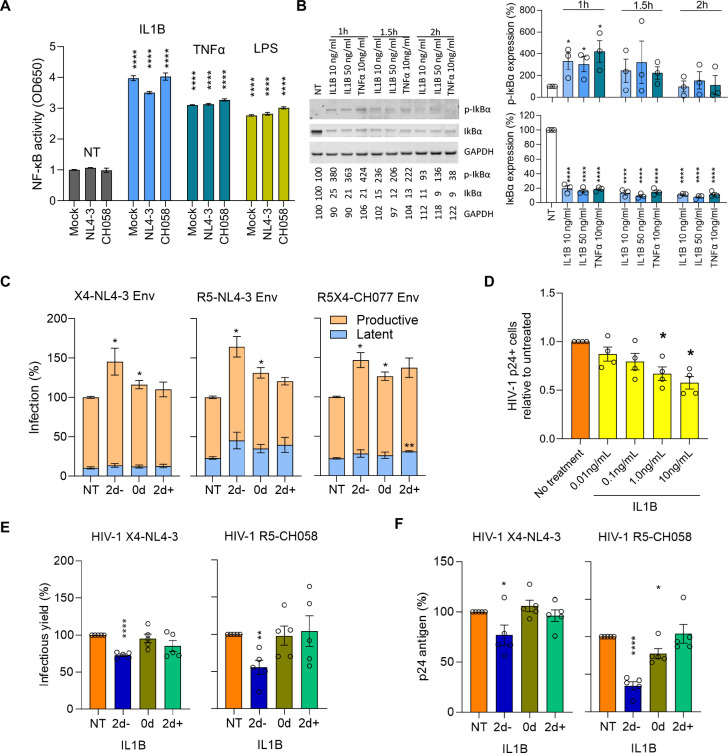
Elimination of latent HIV-1 is a major goal of AIDS research but the host factors determining the size of these reservoirs are poorly understood. Here, we investigated whether differences in host gene expression modulate the size of the HIV-1 reservoir during suppressive ART. Peripheral blood mononuclear cells (PBMC) from fourteen individuals initiating ART during acute infection who demonstrated effective viral suppression but varying magnitude of total HIV-1 DNA were characterized by single-cell RNA sequencing (scRNA-seq). Differentially expressed genes and enriched pathways demonstrated increased monocyte activity in participants with undetectable HIV-1 reservoirs. IL1B expression in CD14+ monocytes showed the greatest fold difference. The inverse association of IL1B with reservoir size was validated in an independent cohort comprised of 38 participants with different genetic backgrounds and HIV-1 subtype infections, and further confirmed with intact proviral DNA assay (IPDA®) measurements of intact HIV-1 proviruses in a subset of the samples. Modeling interactions with cell population frequencies showed that monocyte IL1B expression associated inversely with reservoir size in the context of higher frequencies of central memory CD4+ T cells, implicating an indirect effect of IL1B via the cell type well established to be a reservoir for persistent HIV-1. Signatures consisting of co-expressed genes including IL1B were highly enriched in the "TNFα signaling via NF-κB" geneset. Functional analyses in cell culture revealed that IL1B activates NF-κB, thereby promoting productive HIV-1 infection while simultaneously suppressing viral spread, suggesting a natural latency reversing activity to deplete the reservoir in ART treated individuals. Altogether, unbiased high throughput scRNA-seq analyses revealed that monocyte IL1B variation could decrease HIV-1 proviral reservoirs in individuals initiating ART during acute infection.
Figures
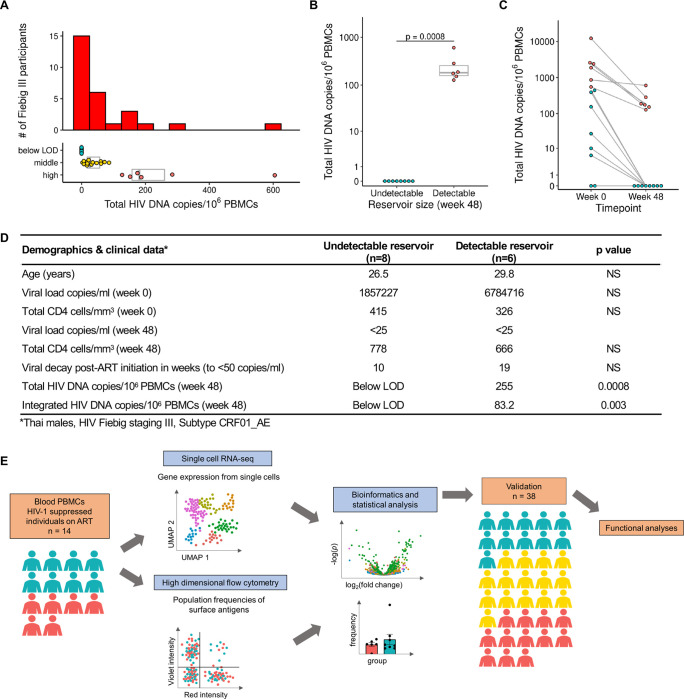
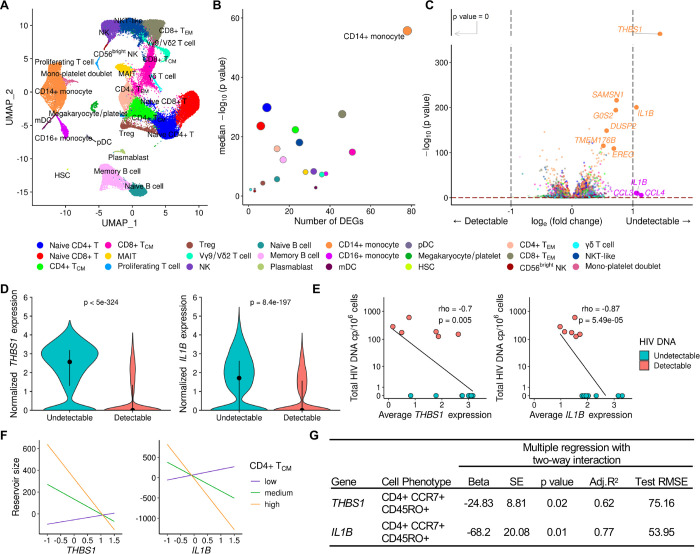
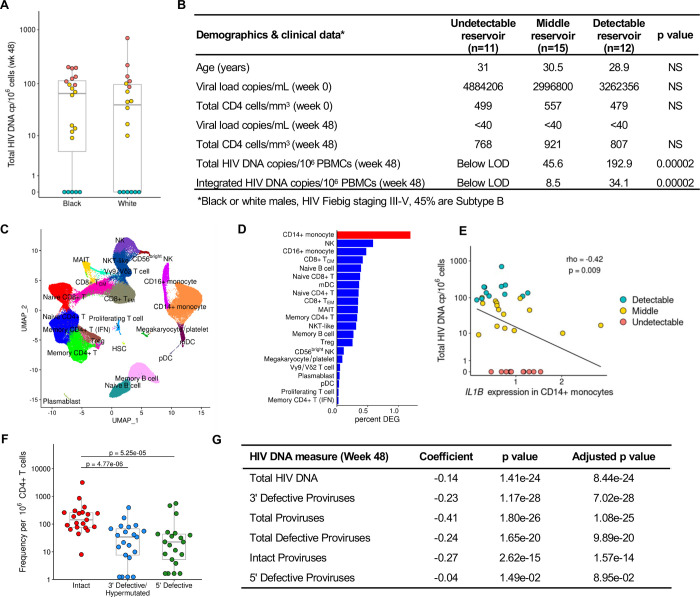
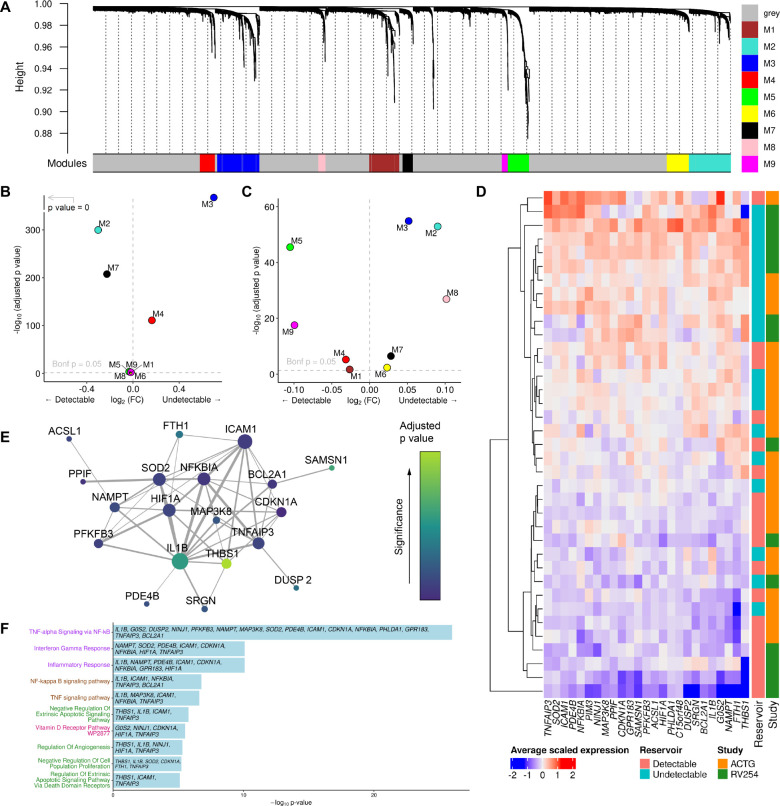
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials