

This is a preprint.
Influenza A virus transcription generates capped cRNAs that activate RIG-I
- PMID: 39605425
- PMCID: PMC11601390
- DOI: 10.1101/2024.11.12.623191
Influenza A virus transcription generates capped cRNAs that activate RIG-I
Abstract
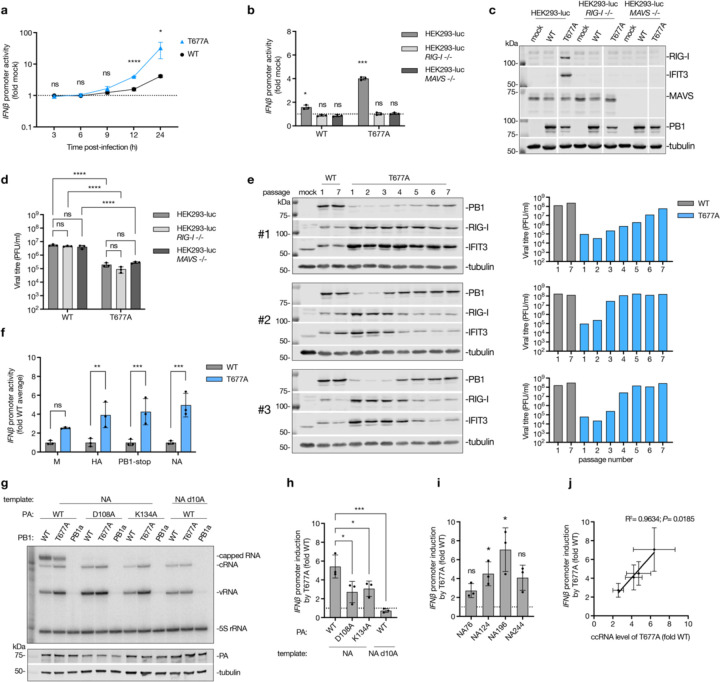
During influenza A virus (IAV) infection, host pathogen receptor retinoic acid-inducible gene I (RIG-I) detects the partially complementary, 5'-triphosphorylated ends of the viral genome segments and non-canonical replication products. However, it has also been suggested that innate immune responses may be triggered by viral transcription. In this study, we investigated whether an immunostimulatory RNA is produced during IAV transcription. We show that the IAV RNA polymerase can read though the polyadenylation signal during transcription termination, generating a capped complementary RNA (ccRNA), which contains the 5' cap of an IAV mRNA and the 3' terminus of a cRNA instead of a poly(A) tail. ccRNAs are detectable in vitro and in both ribonucleoprotein reconstitution assays and IAV infections. Mutations that disrupt polyadenylation enhance ccRNA synthesis and increase RIG-I-dependent innate immune activation. Notably, while ccRNA itself is not immunostimulatory, it forms a RIG-I agonist by hybridizing with a complementary negative-sense viral RNA. These findings thus identify a novel non-canonical IAV RNA species and suggest an alternative mechanism for RIG-I activation during IAV infection.
Keywords: RIG-I; RNA polymerase; aberrant RNA; capped cRNA; influenza A virus; transcription.
Conflict of interest statement
Competing interests The authors declare no competing interests.
Figures



References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous