

This is a preprint.
Electrical synapse molecular diversity revealed by proximity-based proteomic discovery
- PMID: 39605535
- PMCID: PMC11601576
- DOI: 10.1101/2024.11.22.624763
Electrical synapse molecular diversity revealed by proximity-based proteomic discovery
Abstract
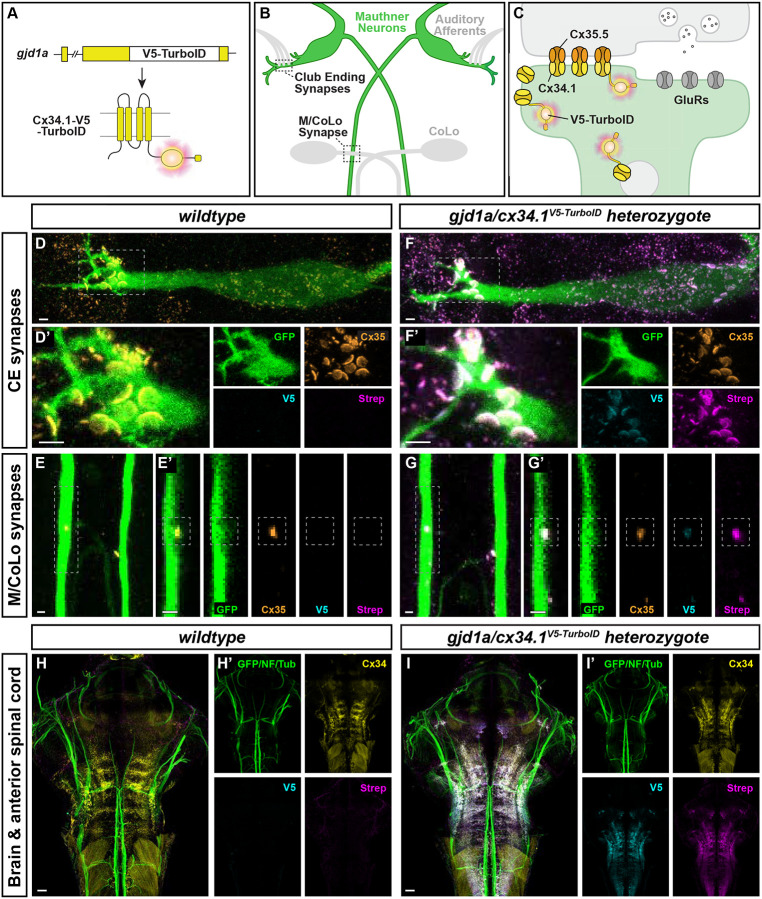
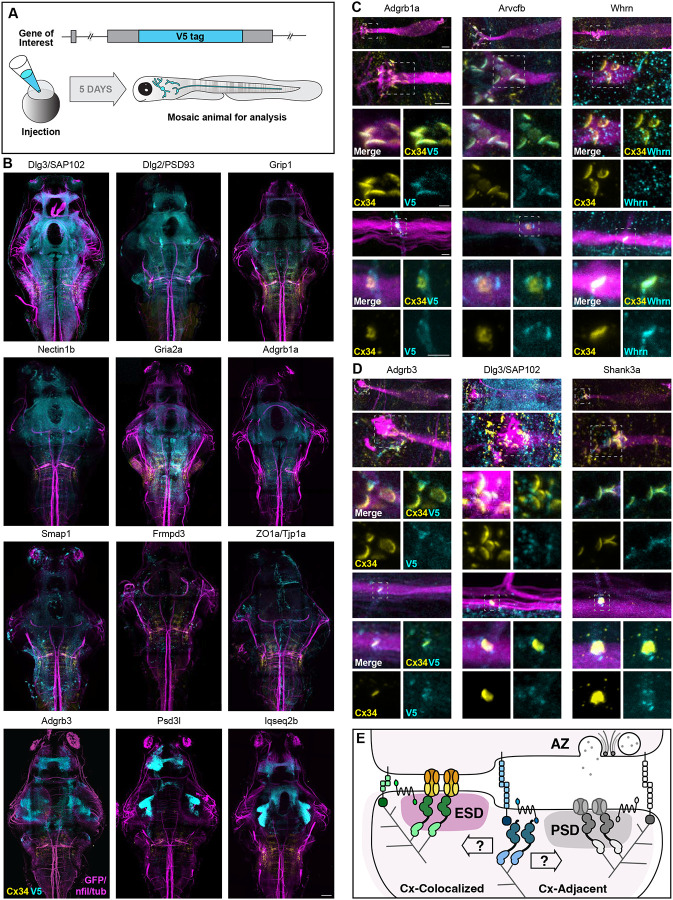
Neuronal circuits are composed of synapses that are either chemical, where signals are transmitted via neurotransmitter release and reception, or electrical, where signals pass directly through interneuronal gap junction channels. While the molecular complexity that controls chemical synapse structure and function is well appreciated, the proteins of electrical synapses beyond the gap-junction-forming Connexins are not well defined. Yet, electrical synapses are expected to be molecularly complex beyond the gap junctions. Connexins are integral membrane proteins requiring vesicular transport and membrane insertion/retrieval to achieve function, homeostasis, and plasticity. Additionally, electron microscopy of neuronal gap junctions reveals neighboring electron dense regions termed the electrical synapse density (ESD). To reveal the molecular complexity of the electrical synapse proteome, we used proximity-dependent biotinylation (TurboID) linked to neural Connexins in zebrafish. Proteomic analysis of developing and mature nervous systems identifies hundreds of Connexin-associated proteins, with overlapping and distinct representation during development and adulthood. The identified protein classes span cell adhesion molecules, cytoplasmic scaffolds, vesicular trafficking, and proteins usually associated with the post synaptic density (PSD) of chemical synapses. Using circuits with stereotyped electrical and chemical synapses, we define molecular sub-synaptic compartments of ESD localizing proteins, we find molecular heterogeneity amongst electrical synapse populations, and we examine the synaptic intermingling of electrical and chemical synapse proteins. Taken together, these results reveal a new complexity of electrical synapse molecular diversity and highlight a novel overlap between chemical and electrical synapse proteomes. Moreover, human homologs of the electrical synapse proteins are associated with autism, epilepsy, and other neurological disorders, providing a novel framework towards understanding neuro-atypical states.
Figures

References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous