

This is a preprint.
APOE Protects Against Severe Infection with Mycobacterium tuberculosis by Restraining Production of Neutrophil Extracellular Traps
- PMID: 39605723
- PMCID: PMC11601580
- DOI: 10.1101/2024.10.04.616580
APOE Protects Against Severe Infection with Mycobacterium tuberculosis by Restraining Production of Neutrophil Extracellular Traps
Update in
-
APOE protects against severe infection with Mycobacterium tuberculosis by restraining production of neutrophil extracellular traps.PLoS Pathog. 2025 Jun 16;21(6):e1013267. doi: 10.1371/journal.ppat.1013267. eCollection 2025 Jun. PLoS Pathog. 2025. PMID: 40523023 Free PMC article.
Abstract
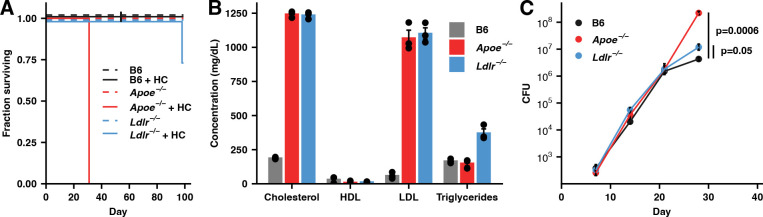
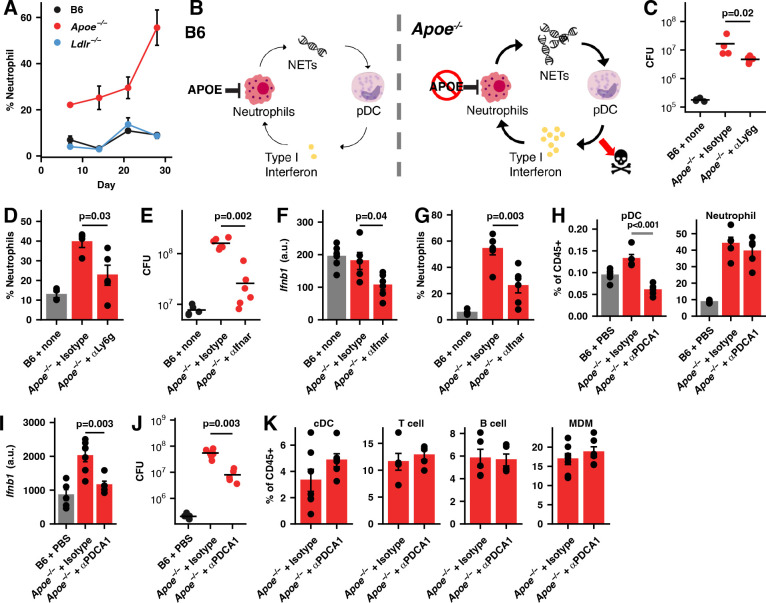
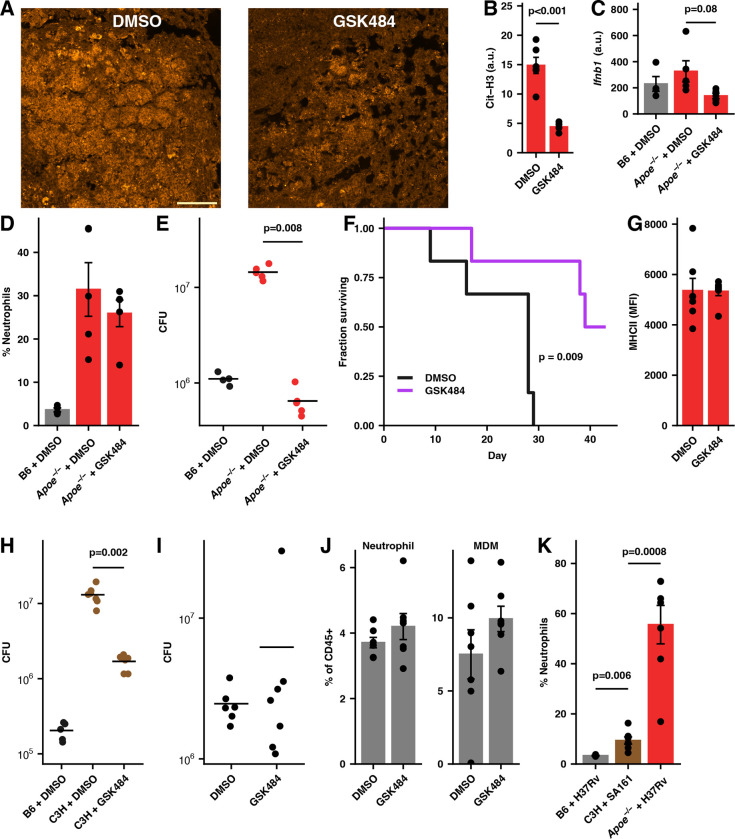
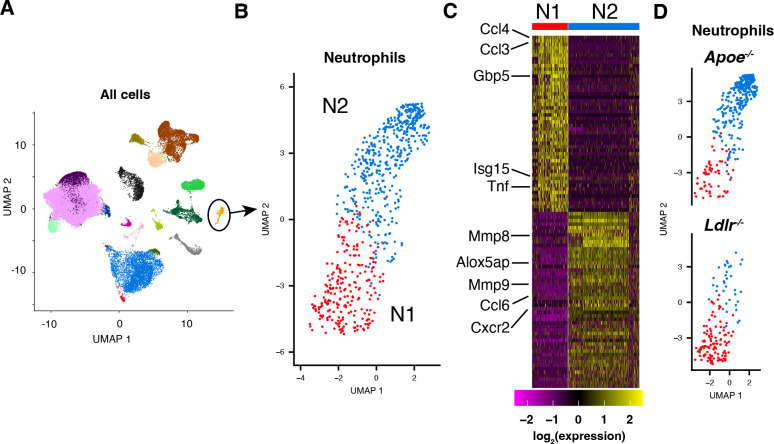
While neutrophils are the predominant cell type in the lungs of humans with active tuberculosis (TB), they are relatively scarce in the lungs of most strains of mice that are used to study the disease. However, similar to humans, neutrophils account for approximately 45% of CD45+ cells in the lungs of Apoe -/- mice on a high-cholesterol (HC) diet following infection with Mycobacterium tuberculosis (Mtb). We hypothesized that the susceptibility of Apoe -/- HC mice might arise from an unrestrained feed-forward loop in which production of neutrophil extracellular traps (NETs) stimulates production of type I interferons by pDCs which in turn leads to the recruitment and activation of more neutrophils, and demonstrated that depleting neutrophils, depleting plasmacytoid dendritic cells (pDCs), or blocking type I interferon signaling, improved the outcome of infection. In concordance with these results, we found that Mtb-infected in Apoe -/- HC mice produce high levels of LTB4 and 12-HETE, two eicosanoids known to act as neutrophil chemoattractants and showed that blocking leukotriene B4 (LTB4) receptor signaling also improved the outcome of tuberculosis. While production of NETs has been associated with severe tuberculosis in other mouse models and in humans, a causative role for NETs in the pathology has not been directly established. We demonstrate that blocking the activation of peptidylarginine deiminase 4 (PAD4), an enzyme critical to NET formation, leads to fewer NETs in the lungs and, strikingly, completely reverses the hypersusceptibility of Apoe -/- HC mice to tuberculosis.
Conflict of interest statement
Competing interests: Authors declare that they have no competing interests.
Figures


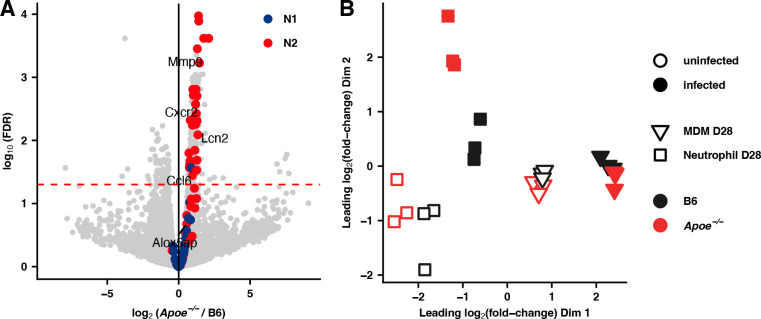
References
-
- Mahley R. W., Apolipoprotein E: cholesterol transport protein with expanding role in cell biology. Science 240, 622–630 (1988). - PubMed
-
- Zhang S. H., Reddick R. L., Piedrahita J. A., Maeda N., Spontaneous hypercholesterolemia and arterial lesions in mice lacking apolipoprotein E. Science 258, 468–471 (1992). - PubMed
-
- Plump A. S., Smith J. D., Hayek T., Aalto-Setälä K., Walsh A., Verstuyft J. G., Rubin E. M., Breslow J. L., Severe hypercholesterolemia and atherosclerosis in apolipoprotein E-deficient mice created by homologous recombination in ES cells. Cell 71, 343–353 (1992). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous