

SPLICER: a highly efficient base editing toolbox that enables in vivo therapeutic exon skipping
- PMID: 39609418
- PMCID: PMC11604662
- DOI: 10.1038/s41467-024-54529-y
SPLICER: a highly efficient base editing toolbox that enables in vivo therapeutic exon skipping
Abstract
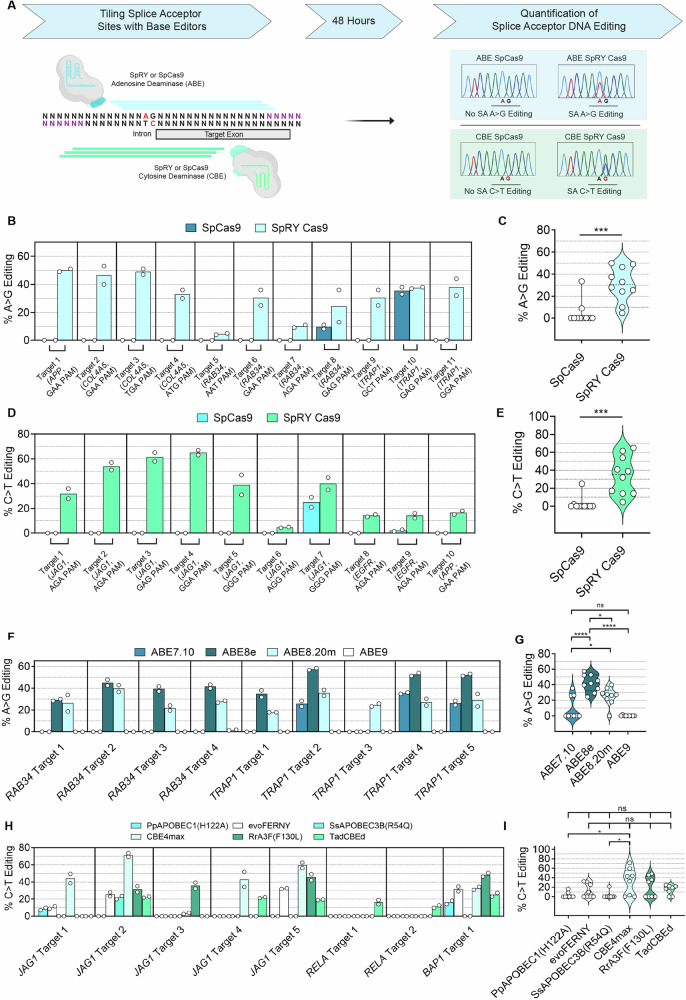
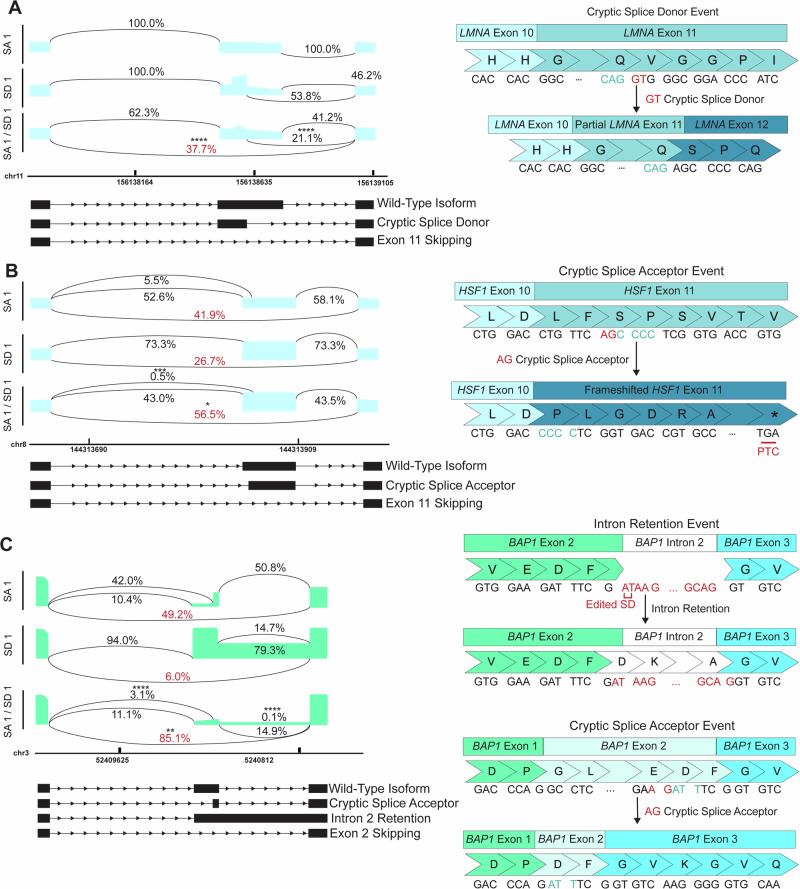
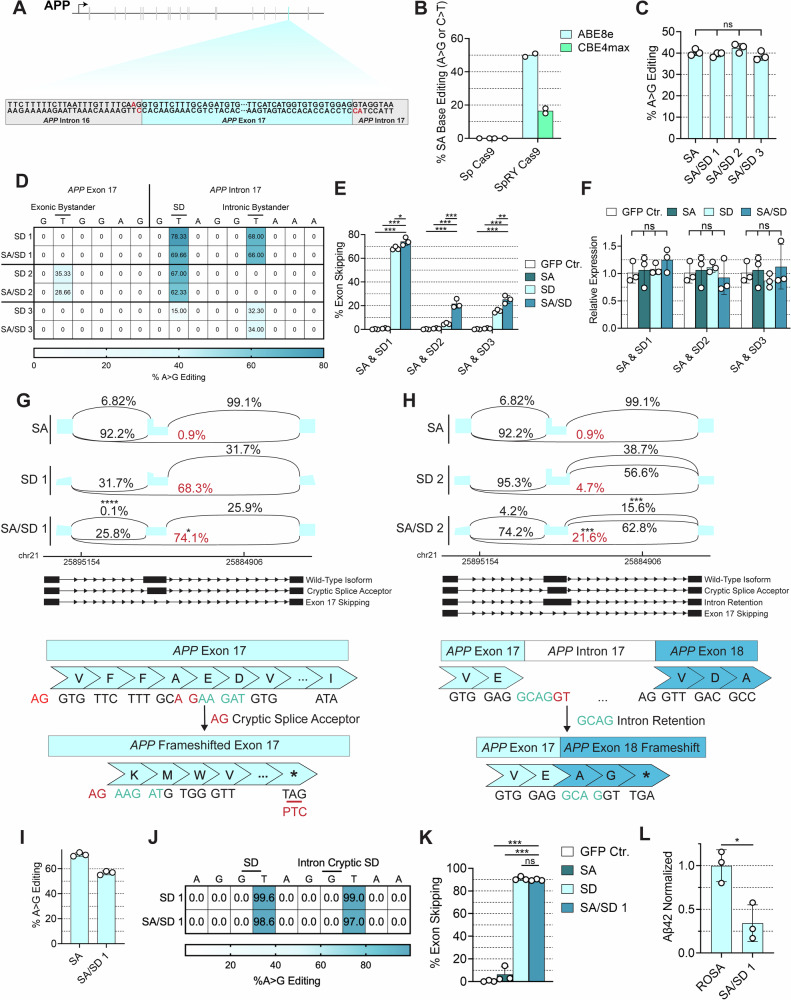
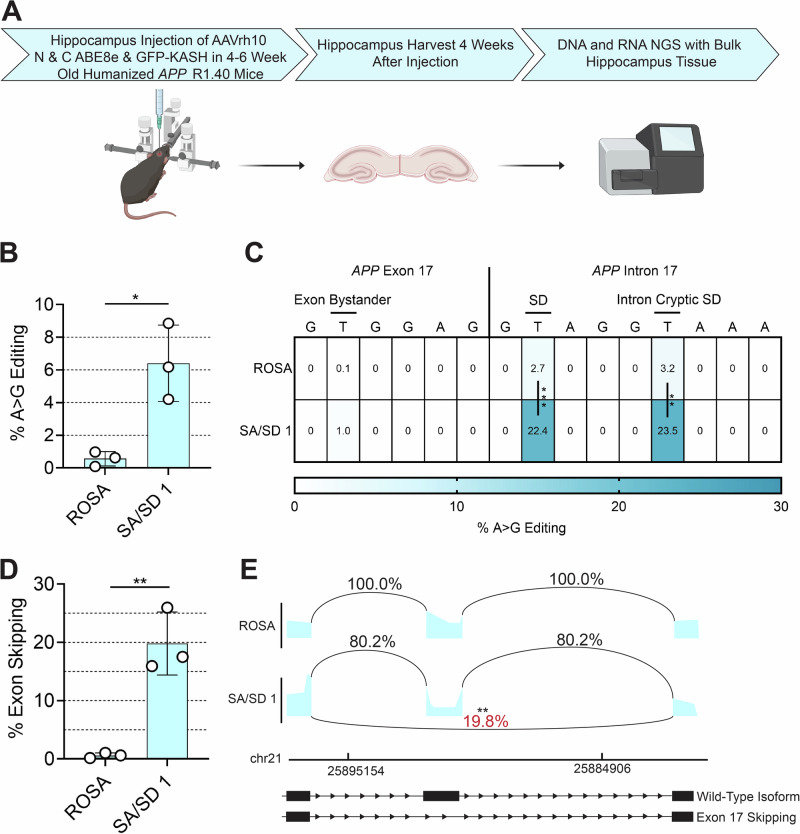
Exon skipping technologies enable exclusion of targeted exons from mature mRNA transcripts, which have broad applications in medicine and biotechnology. Existing techniques including antisense oligonucleotides, targetable nucleases, and base editors, while effective for specific applications, remain hindered by transient effects, genotoxicity, and inconsistent exon skipping. To overcome these limitations, here we develop SPLICER, a toolbox of next-generation base editors containing near-PAMless Cas9 nickase variants fused to adenosine or cytosine deaminases for the simultaneous editing of splice acceptor (SA) and splice donor (SD) sequences. Synchronized SA and SD editing improves exon skipping, reduces aberrant splicing, and enables skipping of exons refractory to single splice site editing. To demonstrate the therapeutic potential of SPLICER, we target APP exon 17, which encodes amino acids that are cleaved to form Aβ plaques in Alzheimer's disease. SPLICER reduces the formation of Aβ42 peptides in vitro and enables efficient exon skipping in a mouse model of Alzheimer's disease. Overall, SPLICER is a widely applicable and efficient exon skipping toolbox.
© 2024. The Author(s).
Conflict of interest statement
Competing interests: P.P.P., M.G., and J.W. are named inventors in pending patent application US17/260,828 filed by the University of Illinois. Methods for exon skipping and AAV-mediated delivery of base editors are covered in the patent application. The remaining authors declare no competing interests.
Figures

Update of
-
SPLICER: A Highly Efficient Base Editing Toolbox That Enables In Vivo Therapeutic Exon Skipping.bioRxiv [Preprint]. 2024 Apr 2:2024.04.01.587650. doi: 10.1101/2024.04.01.587650. bioRxiv. 2024. Update in: Nat Commun. 2024 Nov 28;15(1):10354. doi: 10.1038/s41467-024-54529-y. PMID: 38883727 Free PMC article. Updated. Preprint.
References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
- R01 GM131272/GM/NIGMS NIH HHS/United States
- MDA602798/Muscular Dystrophy Association (Muscular Dystrophy Association Inc.)
- UL1 TR001422/TR/NCATS NIH HHS/United States
- 17SDG33650087/American Heart Association (American Heart Association, Inc.)
- 1U01NS122102/U.S. Department of Health & Human Services | NIH | National Institute of Neurological Disorders and Stroke (NINDS)
- 1R01NS123556/U.S. Department of Health & Human Services | NIH | National Institute of Neurological Disorders and Stroke (NINDS)
- 1R01GM127497/U.S. Department of Health & Human Services | NIH | National Institute of General Medical Sciences (NIGMS)
- R01 GM141296/GM/NIGMS NIH HHS/United States
- 1R01GM141296/U.S. Department of Health & Human Services | NIH | National Institute of General Medical Sciences (NIGMS)
- R01 GM127497/GM/NIGMS NIH HHS/United States
- U01 NS122102/NS/NINDS NIH HHS/United States
- R01 NS123556/NS/NINDS NIH HHS/United States
- T32 EB019944/EB/NIBIB NIH HHS/United States
- 1R01GM131272/U.S. Department of Health & Human Services | NIH | National Institute of General Medical Sciences (NIGMS)
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
