

Functional screening reveals genetic dependencies and diverging cell cycle control in atypical teratoid rhabdoid tumors
- PMID: 39617889
- PMCID: PMC11610224
- DOI: 10.1186/s13059-024-03438-w
Functional screening reveals genetic dependencies and diverging cell cycle control in atypical teratoid rhabdoid tumors
Abstract
Background: Atypical teratoid rhabdoid tumors (ATRT) are incurable high-grade pediatric brain tumors. Despite intensive research efforts, the prognosis for ATRT patients under currently established treatment protocols is poor. While novel therapeutic strategies are urgently needed, the generation of molecular-driven treatment concepts is a challenge mainly due to the absence of actionable genetic alterations.
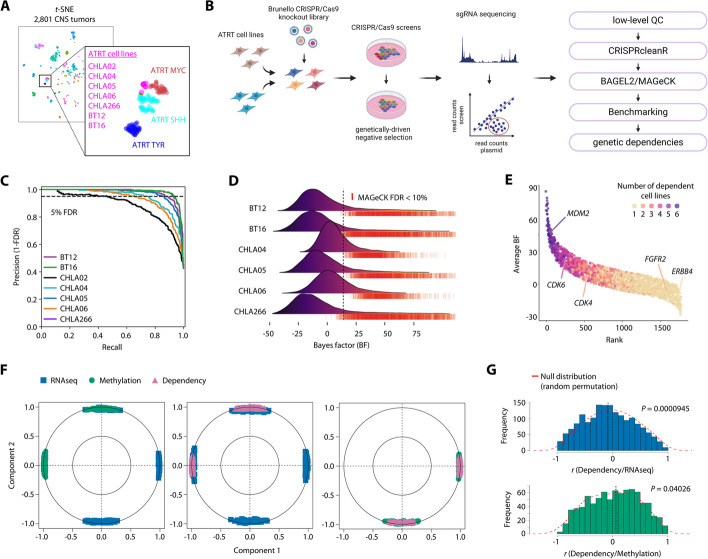
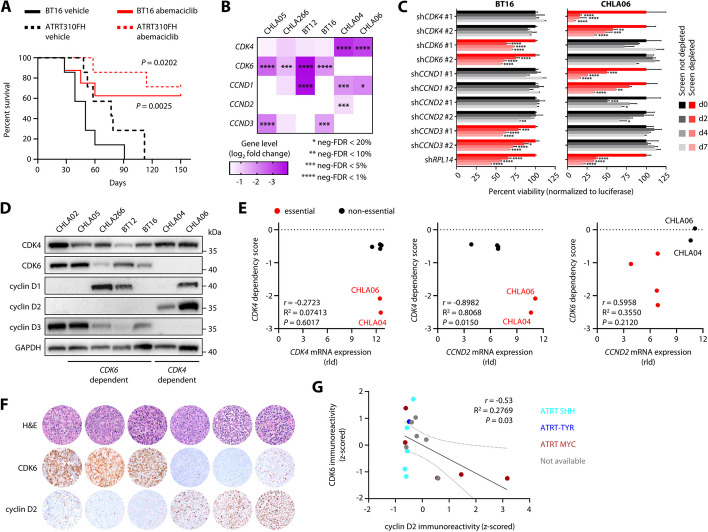
Results: We here use a functional genomics approach to identify genetic dependencies in ATRT, validate selected hits using a functionally instructed small molecule drug library, and observe preferential activity in ATRT cells without subgroup-specific selectivity. CDK4/6 inhibitors are among the most potent drugs and display anti-tumor efficacy due to mutual exclusive dependency on CDK4 or CDK6. Chemogenetic interactor screens reveal a broad spectrum of G1 phase cell cycle regulators that differentially enable cell cycle progression and modulate response to CDK4/6 inhibition in ATRT cells. In this regard, we find that the ubiquitin ligase substrate receptor AMBRA1 acts as a context-specific inhibitor of cell cycle progression by regulating key components of mitosis including aurora kinases.
Conclusions: Our data provide a comprehensive resource of genetic and chemical dependencies in ATRTs, which will inform further preclinical evaluation of novel targeted therapies for this tumor entity. Furthermore, this study reveals a unique mechanism of cell cycle inhibition as the basis for tumor suppressive functions of AMBRA1.
Keywords: AMBRA1; CDK4/6 inhibitors; CRISPR-Cas9; Functional screening; Genetic dependencies; Rhabdoid tumors; Tumor suppressor.
© 2024. The Author(s).
Conflict of interest statement
Declarations. Ethical approval and consent to participate: All animal experiments were approved by the Regierungspräsidium Tübingen (N 09/20 G and N 11/20 M). All experiments were conducted in accordance with the animal law. Animals were closely monitored. For reporting, we followed the ARRIVE guidelines (version 2.0). Consent for publication: Not applicable. Competing interests: GT has served on advisory boards (Bayer, Boehringer Ingelheim, CureVac, Miltenyi Biomedicine, Novocure), as a consultant (Bayer, Boehringer Ingelheim, CureVac), as steering committee member in non-interventional trials (Bayer, Novocure), as a speaker (Novocure, Servier), and financial compensation for all these activities was provided as institutional funding to the University Hospital Tübingen. FP has been affiliated with the Broad Institute while contributing to this study, but is now working at Merck Research Laboratories, Cambridge, USA. This new affiliation does not constitute any financial interest or any other engulfment associated with this study.
Figures






References
-
- Woehrer A, Slavc I, Waldhoer T, Heinzl H, Zielonke N, Czech T, Benesch M, Hainfellner JA, Haberler C. Austrian Brain Tumor R: Incidence of atypical teratoid/rhabdoid tumors in children: a population-based study by the Austrian Brain Tumor Registry, 1996–2006. Cancer. 2010;116:5725–32. - DOI - PubMed
-
- Biegel JA, Tan L, Zhang F, Wainwright L, Russo P, Rorke LB. Alterations of the hSNF5/INI1 gene in central nervous system atypical teratoid/rhabdoid tumors and renal and extrarenal rhabdoid tumors. Clin Cancer Res. 2002;8:3461–7. - PubMed
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
