

Genetically-stable engineered optogenetic gene switches modulate spatial cell morphogenesis in two- and three-dimensional tissue cultures
- PMID: 39622829
- PMCID: PMC11612184
- DOI: 10.1038/s41467-024-54350-7
Genetically-stable engineered optogenetic gene switches modulate spatial cell morphogenesis in two- and three-dimensional tissue cultures
Abstract
Recent advances in tissue engineering have been remarkable, yet the precise control of cellular behavior in 2D and 3D cultures remains challenging. One approach to address this limitation is to genomically engineer optogenetic control of cellular processes into tissues using gene switches that can operate with only a few genomic copies. Here, we implement blue and red light-responsive gene switches to engineer genomically stable two- and three-dimensional mammalian tissue models. Notably, we achieve precise control of cell death and morphogen-directed patterning in 2D and 3D tissues by optogenetically regulating cell necroptosis and synthetic WNT3A signaling at high spatiotemporal resolution. This is accomplished using custom-built patterned LED systems, including digital mirrors and photomasks, as well as laser techniques. These advancements demonstrate the capability of precise spatiotemporal modulation in tissue engineering and open up new avenues for developing programmable 3D tissue and organ models, with significant implications for biomedical research and therapeutic applications.
© 2024. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures

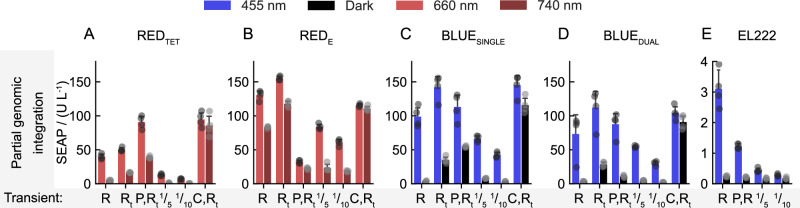



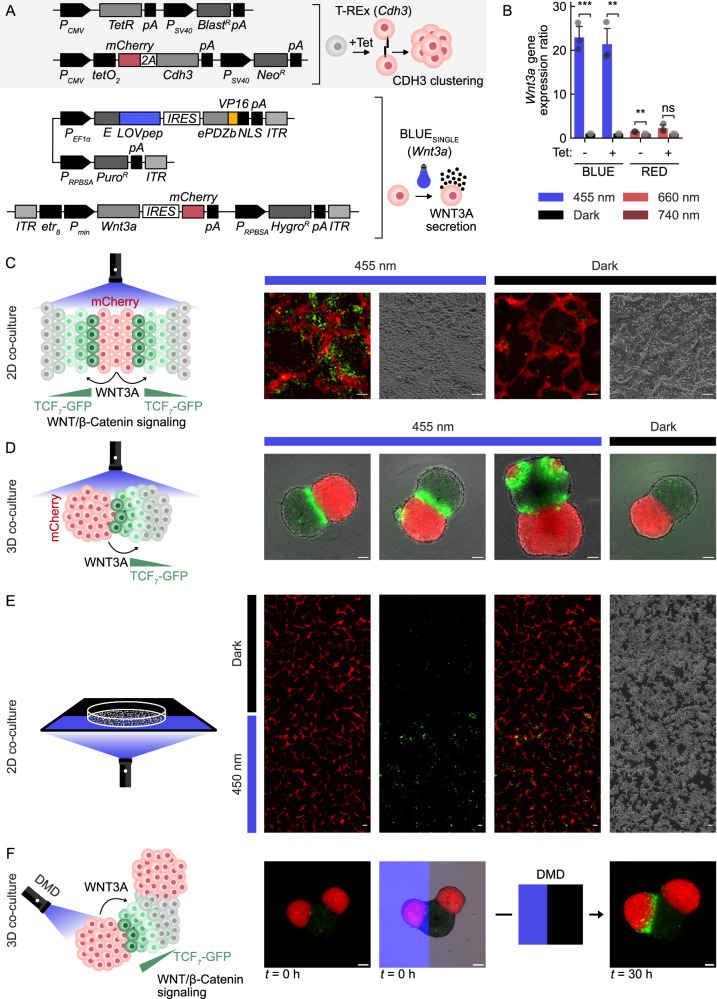
References
-
- Ijäs, T. & Koskinen, R. Exploring biological possibility through synthetic biology. Eur. J. Philos. Sci.11, 39 (2021). - DOI
Publication types
MeSH terms
Substances
Grants and funding
- EXC-2048 Proj no. 390686111/Deutsche Forschungsgemeinschaft (German Research Foundation)
- GRK2466 ID391465903/Deutsche Forschungsgemeinschaft (German Research Foundation)
- CRC1208 ID 267205415 and CRC1535 ID 458090666/Deutsche Forschungsgemeinschaft (German Research Foundation)
- Project ID 801041/EC | EU Framework Programme for Research and Innovation H2020 | H2020 Priority Excellent Science | H2020 Future and Emerging Technologies (H2020 Excellent Science - Future and Emerging Technologies)
- RGY0063/2017 and RGP 0067/2021/Human Frontier Science Program (HFSP)
LinkOut - more resources
Full Text Sources
