

De novo variants in LRRC8C resulting in constitutive channel activation cause a human multisystem disorder
- PMID: 39623139
- PMCID: PMC11729881
- DOI: 10.1038/s44318-024-00322-y
De novo variants in LRRC8C resulting in constitutive channel activation cause a human multisystem disorder
Abstract
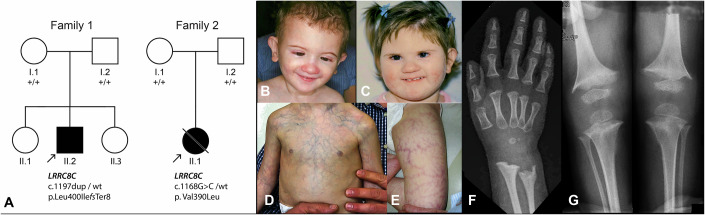
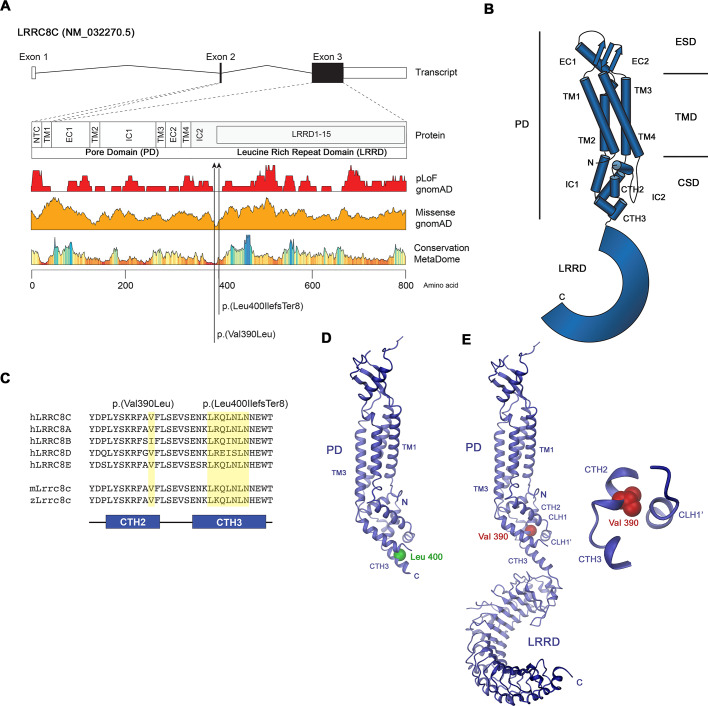
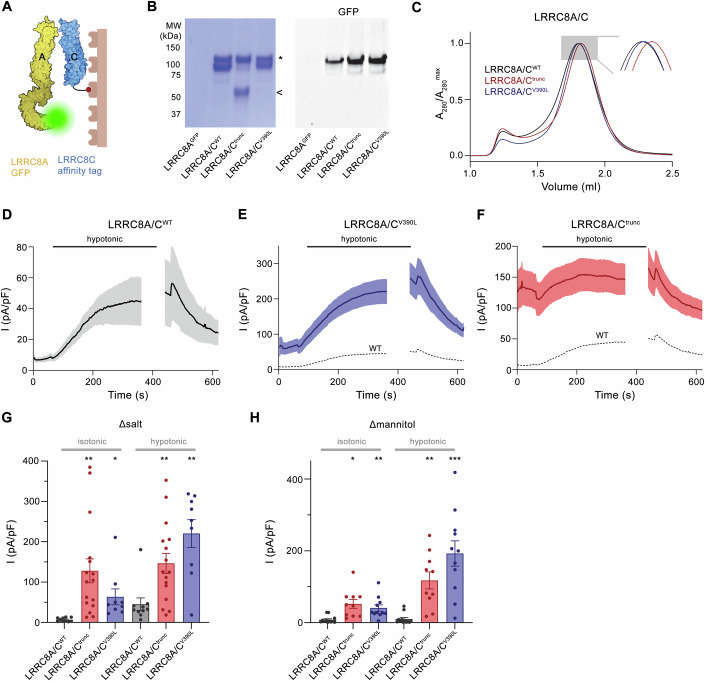
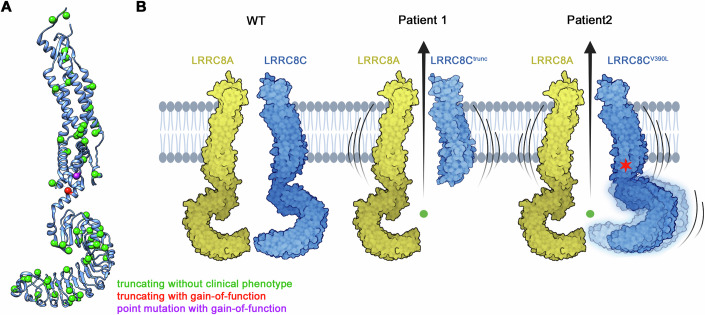
Volume-regulated anion channels (VRACs) are multimeric proteins composed of different paralogs of the LRRC8 family. They are activated in response to hypotonic swelling, but little is known about their specific functions. We studied two human individuals with the same congenital syndrome affecting blood vessels, brain, eyes, and bones. The LRRC8C gene harbored de novo variants in both patients, located in a region of the gene encoding the boundary between the pore and a cytoplasmic domain, which is depleted of sequence variations in control subjects. When studied by cryo-EM, both LRRC8C mutant proteins assembled as their wild-type counterparts, but showed increased flexibility, suggesting a destabilization of subunit interactions. When co-expressed with the obligatory LRRC8A subunit, the mutants exhibited enhanced activation, resulting in channel activity even at isotonic conditions in which wild-type channels are closed. We conclude that structural perturbations of LRRC8C impair channel gating and constitute the mechanistic basis of the dominant gain-of-function effect of these pathogenic variants. The pleiotropic phenotype of this novel clinical entity associated with monoallelic LRRC8C variants indicates the fundamental roles of VRACs in different tissues and organs.
Keywords: Channel Activation; Disease-causing Variants; Volume-regulated Anion Channels.
© 2024. The Author(s).
Conflict of interest statement
Disclosure and competing interests statement. The parents of the two patients in this study have been fully informed that the images shown would be used in a scientific research publication, and that the children will be fully identifiable in this publication, and given their consent. Showing such identifiable patient photographs is necessary for future clinical diagnosis in other affected children of the monoallelic variants of the LRRC8C gene similar to those reported here. We thankfully acknowledge the graciousness and generosity of the parents in giving this consent in the interest of helping future diagnoses in other affected children and in fostering knowledge about and research in this condition. The authors declare no competing interests.
Figures





References
-
- Best L, Brown PD, Sener A, Malaisse WJ (2010) Electrical activity in pancreatic islet cells: The VRAC hypothesis. Islets 2:59–64 - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
