

Epi-microRNA mediated metabolic reprogramming counteracts hypoxia to preserve affinity maturation
- PMID: 39627218
- PMCID: PMC11615350
- DOI: 10.1038/s41467-024-54937-0
Epi-microRNA mediated metabolic reprogramming counteracts hypoxia to preserve affinity maturation
Abstract
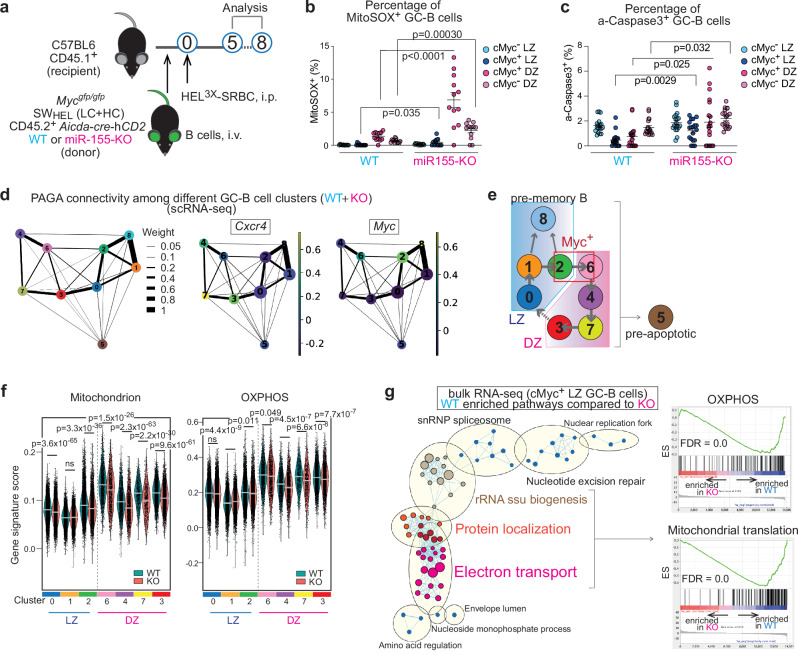
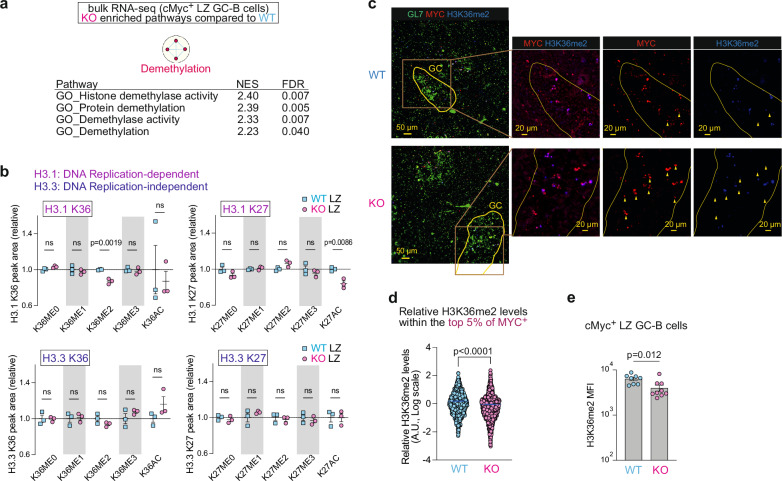
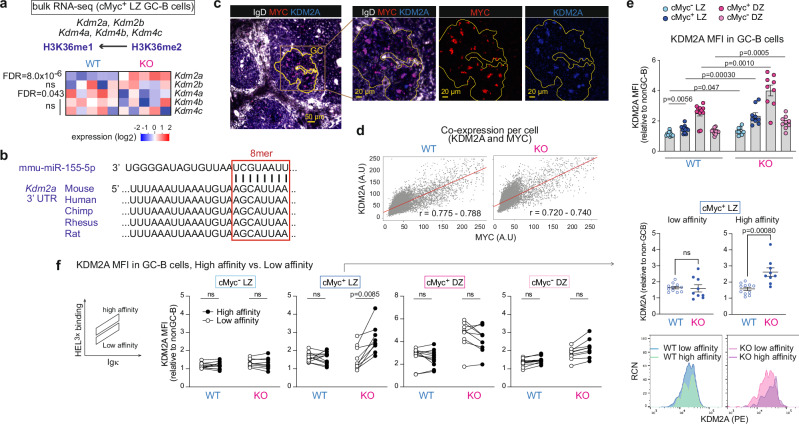
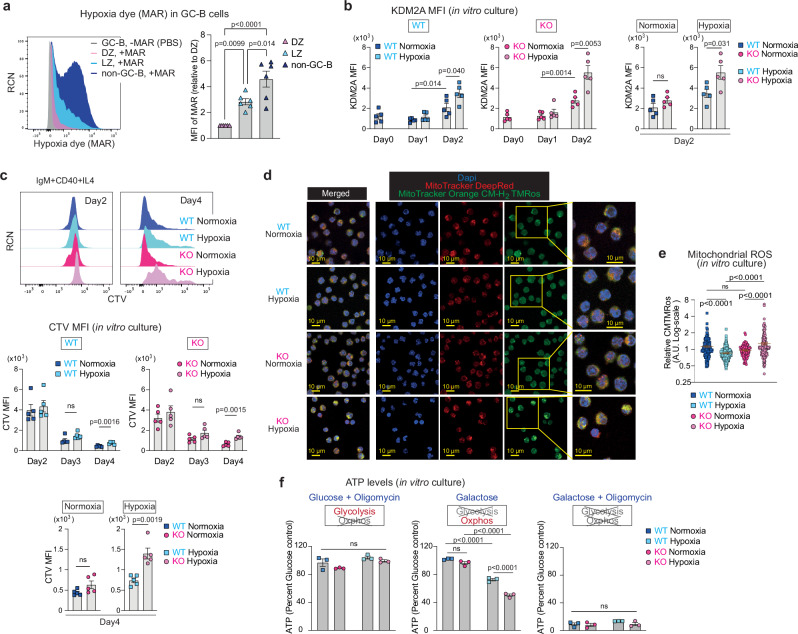
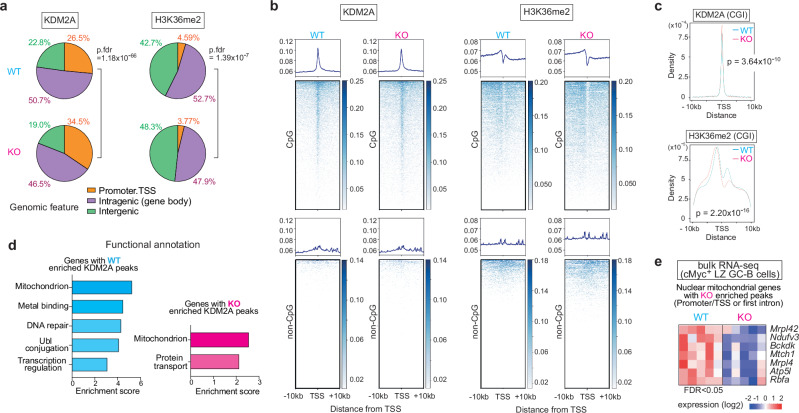
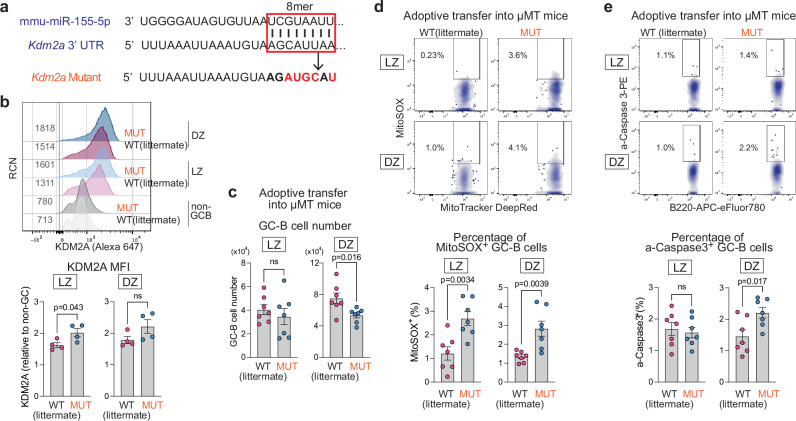
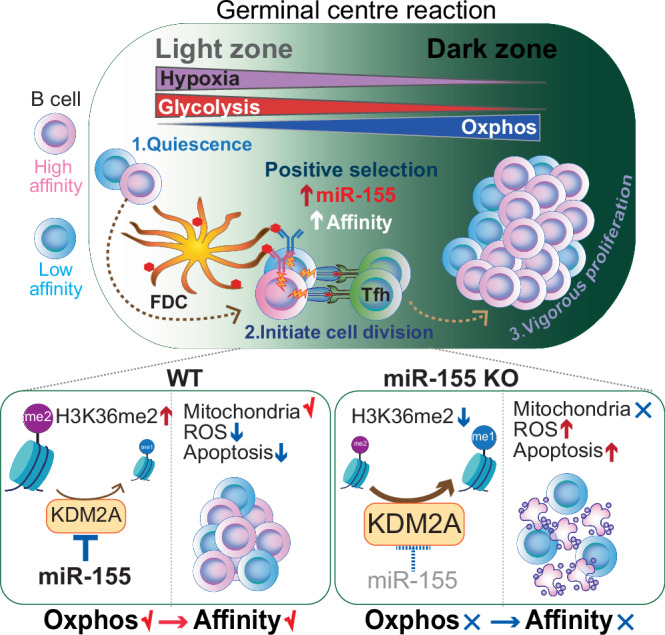
To increase antibody affinity against pathogens, positively selected GC-B cells initiate cell division in the light zone (LZ) of germinal centers (GCs). Among these, higher-affinity clones migrate to the dark zone (DZ) and vigorously proliferate by utilizing energy provided by oxidative phosphorylation (OXPHOS). However, it remains unknown how positively selected GC-B cells adapt their metabolism for cell division in the glycolysis-dominant, cell cycle arrest-inducing, hypoxic LZ microenvironment. Here, we show that microRNA (miR)-155 mediates metabolic reprogramming during positive selection to protect high-affinity clones. Mechanistically, miR-155 regulates H3K36me2 levels in hypoxic conditions by directly repressing the histone lysine demethylase, Kdm2a, whose expression increases in response to hypoxia. The miR-155-Kdm2a interaction is crucial for enhancing OXPHOS through optimizing the expression of vital nuclear mitochondrial genes under hypoxia, thereby preventing excessive production of reactive oxygen species and subsequent apoptosis. Thus, miR-155-mediated epigenetic regulation promotes mitochondrial fitness in high-affinity GC-B cells, ensuring their expansion and consequently affinity maturation.
© 2024. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no direct competing financial or nonfinancial interests.
Figures
Update of
-
Epi-microRNA mediated metabolic reprogramming ensures affinity maturation.bioRxiv [Preprint]. 2023 Oct 31:2023.07.31.551250. doi: 10.1101/2023.07.31.551250. bioRxiv. 2023. Update in: Nat Commun. 2024 Dec 3;15(1):10516. doi: 10.1038/s41467-024-54937-0. PMID: 37609190 Free PMC article. Updated. Preprint.
References
-
- Berek, C., Berger, A. & Apel, M. Maturation of the immune response in germinal centers. Cell67, 1121–1129 (1991). - PubMed
-
- Jacob, J., Kelsoe, G., Rajewsky, K. & Weiss, U. Intraclonal generation of antibody mutants in germinal centres. Nature354, 389–392 (1991). - PubMed
-
- Kepler, T. B. & Perelson, A. S. Cyclic re-entry of germinal center B cells and the efficiency of affinity maturation. Immunol. Today14, 412–415 (1993). - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
