

Repeat-induced point mutations driving Parastagonospora nodorum genomic diversity are balanced by selection against non-synonymous mutations
- PMID: 39627497
- PMCID: PMC11615325
- DOI: 10.1038/s42003-024-07327-7
Repeat-induced point mutations driving Parastagonospora nodorum genomic diversity are balanced by selection against non-synonymous mutations
Abstract
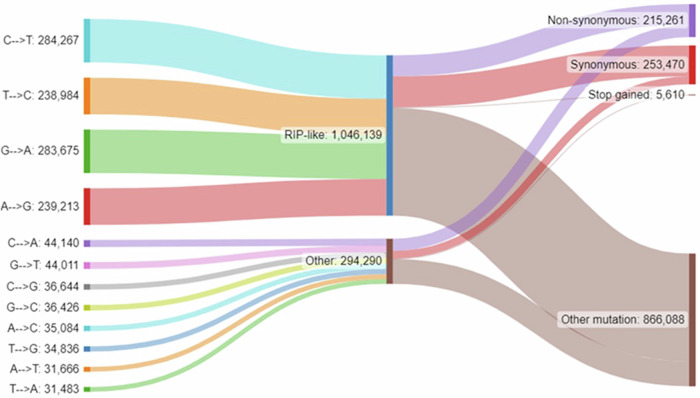
Parastagonospora nodorum is necrotrophic fungal pathogen of wheat with significant genomic resources. Population-level pangenome data for 173 isolates, of which 156 were from Western Australia (WA) and 17 were international, were examined for overall genomic diversity and effector gene content. A heterothallic core population occurred across all regions of WA, with asexually-reproducing clonal clusters in dryer northern regions. High potential for SNP diversity in the form of repeat-induced point mutation (RIP)-like transitions, was observed across the genome, suggesting widespread 'RIP-leakage' from transposon-rich repetitive sequences into non-repetitive regions. The strong potential for RIP-like mutations was balanced by negative selection against non-synonymous SNPs, that was observed within protein-coding regions. Protein isoform profiles of known effector loci (SnToxA, SnTox1, SnTox3, SnTox267, and SnTox5) indicated low-levels of non-synonymous and high-levels of silent RIP-like mutations. Effector predictions identified 186 candidate secreted predicted effector proteins (CSEPs), 69 of which had functional annotations and included confirmed effectors. Pangenome-based effector isoform profiles across WA were distinct from global isolates and were conserved relative to population structure, and may enable new approaches for monitoring crop disease pathotypes.
© 2024. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures



References
-
- Solomon, P. S., Lowe, R. G. T., Tan, K.-C., Waters, O. D. C. & Oliver, R. P. Stagonospora nodorum: cause of stagonospora nodorum blotch of wheat. Mol. Plant Pathol.7, 147–156 (2006). - PubMed
-
- Murray, G. M. & Brennan, J. P. Estimating disease losses to the Australian wheat industry. Austral. Plant Pathol.38, 558–570 (2009).
-
- McDonald, M. C., Razavi, M., Friesen, T. L., Brunner, P. C. & McDonald, B. A. Phylogenetic and population genetic analyses of Phaeosphaeria nodorum and its close relatives indicate cryptic species and an origin in the Fertile Crescent. Fungal Genet. Biol.49, 882–895 (2012). - PubMed
-
- Stukenbrock, E. H., Banke, S. & McDonald, B. A. Global migration patterns in the fungal wheat pathogen Phaeosphaeria nodorum. Mol. Ecol.15, 2895–2904 (2006). - PubMed
-
- Bathgate, J. A. & Loughman, R. Ascospores are a source of inoculum of Phaeosphaeria nodorum, P. avenaria f. sp. avenaria and Mycosphaerella graminicola in Western Australia. Austral. Plant Pathol.30, 317 (2001).
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
