

Circuit mechanism underlying fragmented sleep and memory deficits in 16p11.2 deletion mouse model of autism
- PMID: 39628570
- PMCID: PMC11612818
- DOI: 10.1016/j.isci.2024.111285
Circuit mechanism underlying fragmented sleep and memory deficits in 16p11.2 deletion mouse model of autism
Abstract
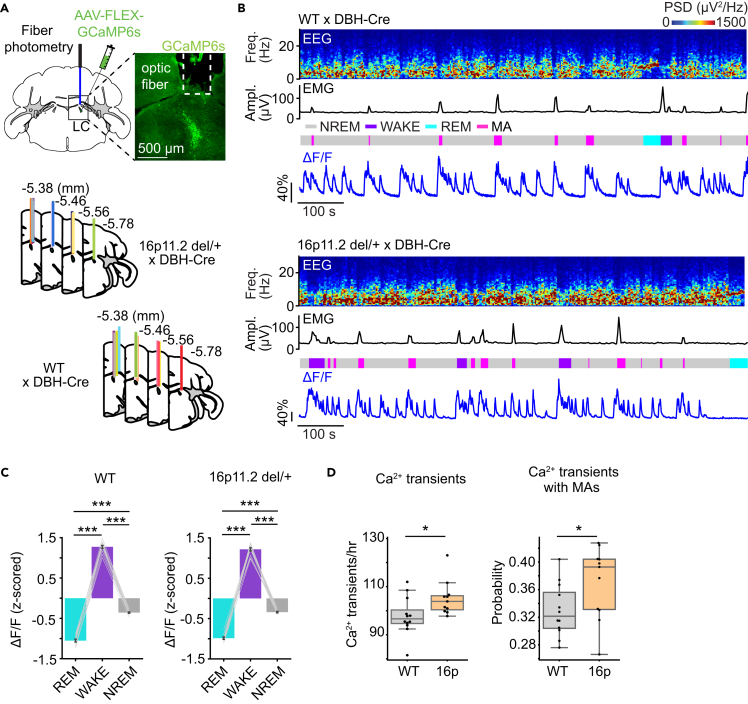
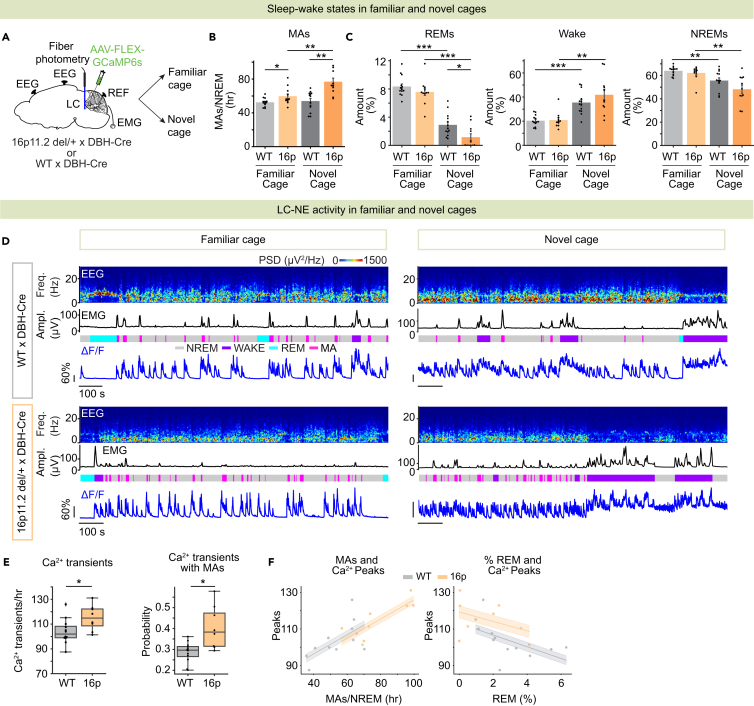
Sleep disturbances are prevalent in children with autism spectrum disorder (ASD). Strikingly, sleep problems are positively correlated with the severity of ASD symptoms, such as memory impairment. However, the neural mechanisms underlying sleep disturbances and cognitive deficits in ASD are largely unexplored. Here, we show that non-rapid eye movement sleep (NREMs) is fragmented in the 16p11.2 deletion mouse model of ASD. The degree of sleep fragmentation is reflected in an increased number of calcium transients in the activity of locus coeruleus noradrenergic (LC-NE) neurons during NREMs. In contrast, optogenetic inhibition of LC-NE neurons and pharmacological blockade of noradrenergic transmission using clonidine consolidate sleep. Furthermore, inhibiting LC-NE neurons restores memory. Finally, rabies-mediated screening of presynaptic neurons reveals altered connectivity of LC-NE neurons with sleep- and memory-regulatory regions in 16p11.2 deletion mice. Our findings identify a crucial role of the LC-NE system in regulating sleep stability and memory in ASD.
Keywords: Behavioral neuroscience; Cellular neuroscience; Molecular neuroscience.
© 2024 The Authors.
Conflict of interest statement
The authors declare no competing financial interests.
Figures





Update of
-
Circuit mechanism underlying fragmented sleep and memory deficits in 16p11.2 deletion mouse model of autism.bioRxiv [Preprint]. 2024 May 21:2023.12.26.573156. doi: 10.1101/2023.12.26.573156. bioRxiv. 2024. Update in: iScience. 2024 Oct 28;27(12):111285. doi: 10.1016/j.isci.2024.111285. PMID: 38234815 Free PMC article. Updated. Preprint.
-
Circuit mechanism underlying fragmented sleep and memory deficits in 16p11.2 deletion mouse model of autism.Res Sq [Preprint]. 2024 Mar 14:rs.3.rs-3877710. doi: 10.21203/rs.3.rs-3877710/v1. Res Sq. 2024. Update in: iScience. 2024 Oct 28;27(12):111285. doi: 10.1016/j.isci.2024.111285. PMID: 38559267 Free PMC article. Updated. Preprint.
References
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
