

CRISPR-based gene editing technology and its application in microbial engineering
- PMID: 39628916
- PMCID: PMC11610974
- DOI: 10.1016/j.engmic.2023.100101
CRISPR-based gene editing technology and its application in microbial engineering
Abstract
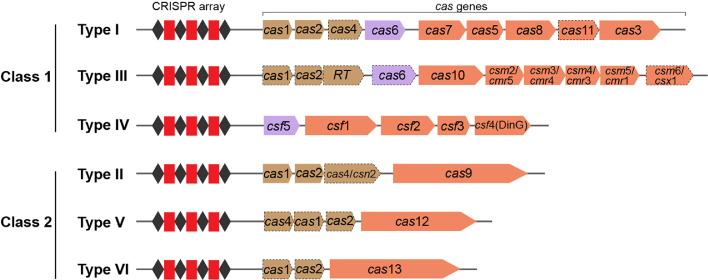
Gene editing technology involves the modification of a specific target gene to obtain a new function or phenotype. Recent advances in clustered regularly interspaced short palindromic repeats (CRISPR)-Cas-mediated technologies have provided an efficient tool for genetic engineering of cells and organisms. Here, we review the three emerging gene editing tools (ZFNs, TALENs, and CRISPR-Cas) and briefly introduce the principle, classification, and mechanisms of the CRISPR-Cas systems. Strategies for gene editing based on endogenous and exogenous CRISPR-Cas systems, as well as the novel base editor (BE), prime editor (PE), and CRISPR-associated transposase (CAST) technologies, are described in detail. In addition, we summarize recent developments in the application of CRISPR-based gene editing tools for industrial microorganism and probiotics modifications. Finally, the potential challenges and future perspectives of CRISPR-based gene editing tools are discussed.
Keywords: CRISPR-Cas system; Gene editing; Industrial microorganisms; Probiotics.
© 2023 The Author(s).
Conflict of interest statement
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures





References
-
- Jasin M. Genetic manipulation of genomes with rare-cutting endonucleases. Trends Genet. TIG. 1996;12:224–228. - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Miscellaneous
