

Phosphatidylinositol 4,5-bisphosphate drives the formation of EGFR and EphA2 complexes
- PMID: 39630914
- PMCID: PMC11616708
- DOI: 10.1126/sciadv.adl0649
Phosphatidylinositol 4,5-bisphosphate drives the formation of EGFR and EphA2 complexes
Abstract
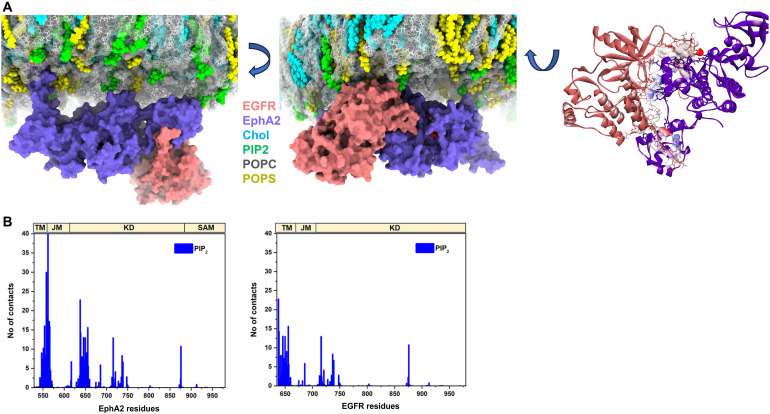
Receptor tyrosine kinases (RTKs) regulate many cellular functions and are important targets in pharmaceutical development, particularly in cancer treatment. EGFR and EphA2 are two key RTKs that are associated with oncogenic phenotypes. Several studies have reported functional interplay between these receptors, but the mechanism of interaction is still unresolved. Here, we use a time-resolved fluorescence spectroscopy called PIE-FCCS to resolve EGFR and EphA2 interactions in live cells. We tested the role of ligands and found that EGF, but not ephrin A1 (EA1), stimulated heteromultimerization between the receptors. To determine the effect of anionic lipids, we targeted phospholipase C (PLC) activity to alter the abundance of phosphatidylinositol 4,5-bisphosphate (PIP2). We found that higher PIP2 levels increased homomultimerization of both EGFR and EphA2, as well as heteromultimerization. This study provides a direct characterization of EGFR and EphA2 interactions in live cells and shows that PIP2 can have a substantial effect on the spatial organization of RTKs.
Figures






Update of
-
Phosphatidylinositol (4,5)-bisphosphate drives the formation of EGFR and EphA2 complexes.bioRxiv [Preprint]. 2024 May 4:2024.05.03.592400. doi: 10.1101/2024.05.03.592400. bioRxiv. 2024. Update in: Sci Adv. 2024 Dec 6;10(49):eadl0649. doi: 10.1126/sciadv.adl0649. PMID: 38746348 Free PMC article. Updated. Preprint.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
