

Sequence of formation and inheritance of meristic variation in the post-cranial axial skeleton of Atlantic salmon (Salmo salar)
- PMID: 39631427
- PMCID: PMC11949744
- DOI: 10.1111/jfb.16004
Sequence of formation and inheritance of meristic variation in the post-cranial axial skeleton of Atlantic salmon (Salmo salar)
Abstract
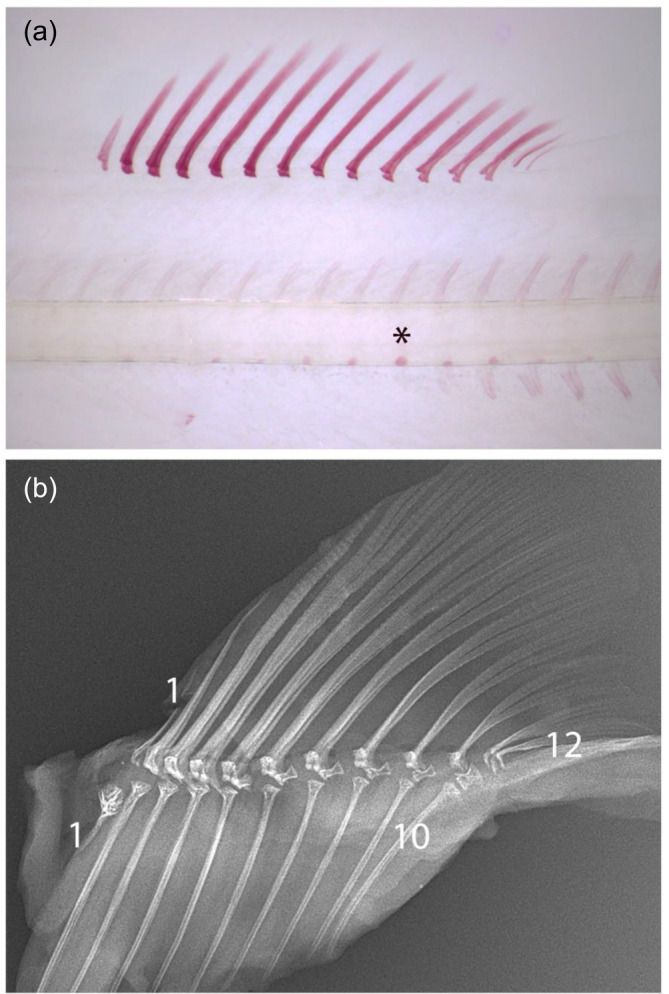
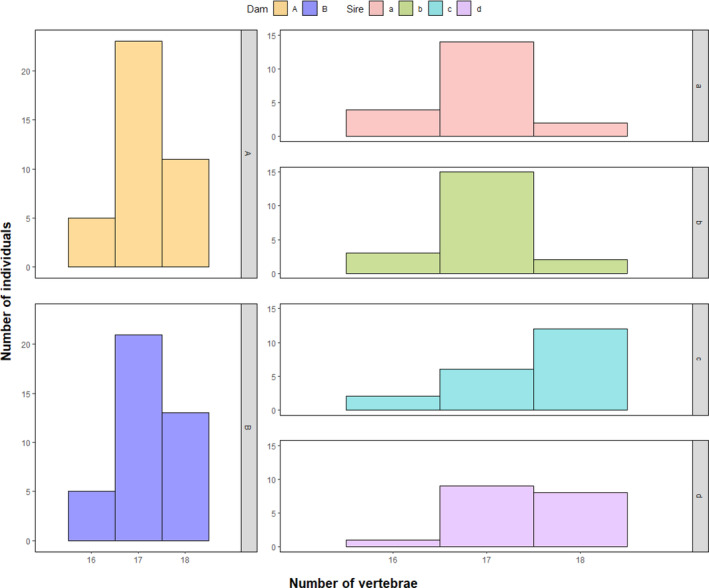
Atlantic salmon is an important aquaculture species that has fascinated naturalists for centuries, resulting in its biology being widely characterized. Certain details about the early development and the inheritance of meristic variation in the post-cranial axial skeleton are, however, largely unexplored. The present study gives a detailed description of the sequence of formation of the post-cranial axial skeleton based on whole-mount staining and used radiology to investigate the inheritance of meristic variation in isogenic hybrid all-male families of Atlantic salmon (~4 kg). Eight different families were created by crossing two homozygous double haploid XX females (dam A, B) with four different double haploid homozygous YY super males (sires a to d). In the caudal fin complex, the first bone to form is hypural 1 and its associated lepidotrichia followed by a bidirectional formation of new bones. In the dorsal and anal fins, development starts in the cranial part, and new bones form bidirectionally towards the head and tail fin. The neural and haemal arches start to form at segment 43, and further development is bidirectional. The first parapophysis form in the caudal part of the abdomen followed by a unidirectional completion cranially. The first ribs form at segment 3 and new ribs develop unidirectional caudally. Chordacentra formation starts at segment 24 followed by formation of chordacentrum number 58 (caudal-most vertebra). New chordacentrae form bidirectionally from segment 24 in parallel with the formation of chordacentrum number 57. The first epineuralia form at segment 1 followed by a unidirectional completion caudally until segment 30. The first supraneuralia to develop is number 10 closely followed by number 1, then new supraneurals form bidirectionally from number 10. Analysis of the inheritance on the post-cranial axial skeletal bones showed a strong maternal effect on total vertebrae centra and tail fin lepidotrichia counts. For these skeletal counts, dam A produced offspring with modes of 58 and 45 respectively, while dam B produced offspring with modes of 59 and 42. The higher number of total vertebrae centra produced by dam B was associated with additional abdominal and/or transitional vertebrae. The completion of formation in different post-cranial axial skeletal parts are either bi- or unidirectional, and the initiation of formation is site specific for each skeletal part with some inter-part similarities. Further, the present results may suggest that there has been a maternally driven selection for more abdominal vertebrae associated with a higher number of total vertebrae, and more tail fin lepidotrichia associated with a lower number of total vertebrae. These changing meristic counts may impact on important fitness-related traits, such as fecundity and swimming ability, making the present findings relevant for both ecological and aquaculture sciences.
Keywords: Atlantic salmon; inheritance; lepidotrich; meristic; pterygiophore; vertebra.
© 2024 The Author(s). Journal of Fish Biology published by John Wiley & Sons Ltd on behalf of Fisheries Society of the British Isles.
Figures









Similar articles
-
Effect of heterozygosity, ploidy and incubation temperature on post-cranial axial skeletal meristics and deformities in Atlantic salmon (Salmo salar).J Fish Biol. 2025 Aug;107(2):441-453. doi: 10.1111/jfb.70032. Epub 2025 Apr 9. J Fish Biol. 2025. PMID: 40200847 Free PMC article.
-
Reevaluation of the caudal skeleton of certain actinopterygian fishes: III. Salmonidae. Homologization of caudal skeletal structures.J Morphol. 1992 Nov;214(2):187-249. doi: 10.1002/jmor.1052140209. J Morphol. 1992. PMID: 29865606
-
Heads and tails: The notochord develops differently in the cranium and caudal fin of Atlantic Salmon (Salmo salar, L.).Anat Rec (Hoboken). 2021 Aug;304(8):1629-1649. doi: 10.1002/ar.24562. Epub 2020 Nov 18. Anat Rec (Hoboken). 2021. PMID: 33155751 Free PMC article.
-
A critical review of adaptive genetic variation in Atlantic salmon: implications for conservation.Biol Rev Camb Philos Soc. 2007 May;82(2):173-211. doi: 10.1111/j.1469-185X.2006.00004.x. Biol Rev Camb Philos Soc. 2007. PMID: 17437557 Review.
-
Atlantic salmon (Salmo salar L.) genetics in the 21st century: taking leaps forward in aquaculture and biological understanding.Anim Genet. 2019 Feb;50(1):3-14. doi: 10.1111/age.12748. Epub 2018 Nov 14. Anim Genet. 2019. PMID: 30426521 Free PMC article. Review.
Cited by
-
Effect of heterozygosity, ploidy and incubation temperature on post-cranial axial skeletal meristics and deformities in Atlantic salmon (Salmo salar).J Fish Biol. 2025 Aug;107(2):441-453. doi: 10.1111/jfb.70032. Epub 2025 Apr 9. J Fish Biol. 2025. PMID: 40200847 Free PMC article.
References
-
- Aguirre, W. E. , Young, A. , Navarrete‐Amaya, R. , Valdiviezo‐Rivera, J. , Jiménez‐Prado, P. , Cucalón, R. V. , Nugra‐Salazar, F. , Calle‐Delgado, P. , Borders, T. , & Shervette, V. R. (2019). Vertebral number covaries with body form and elevation along the western slopes of the Ecuadorian Andes in the Neotropical fish genus Rhoadsia (Teleostei: Characidae). Biological Journal of the Linnean Society, 126, 706–720.
-
- Amos, M. H. , Anas, R. E. , & Pearson, R. E. (1960). Use of a discriminant function in the morphological separation of Asian and north American races of pink salmon, Oncorhynchus gorbuscha (Walbaum). Bureau of Commercial Fisheries.
-
- Ando, D. , Mano, S. I. , Koide, N. , & Nakajima, M. (2008). Estimation of heritability and genetic correlation of number of abdominal and caudal vertebrae in masu salmon. Fisheries Science, 74, 293–298.
-
- Ando, D. , Murooka, M. , Shimoda, K. , Hayano, H. , Sasaki, Y. , Miyakoshi, Y. , & Nakajima, M. (2017). Estimation of heritability of vertebral number in chum salmon Oncorhynchus keta . Fisheries Science, 83, 413–423.
-
- Ando, D. , Shimado, K. , Takeuchi, K. , Iijima, A. , Urabe, H. , Shinriki, Y. , & Nakajima, M. (2019). Comparison of vertebral number between wild and hatchery‐reared population, and effect of stock enhancement programs on vertebral number in masu salmon Oncorhynchus masou . Nippon Suisan Gakkaishi, 85, 487–493.
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources