

A landscape of X-inactivation during human T cell development
- PMID: 39632794
- PMCID: PMC11618795
- DOI: 10.1038/s41467-024-54110-7
A landscape of X-inactivation during human T cell development
Abstract
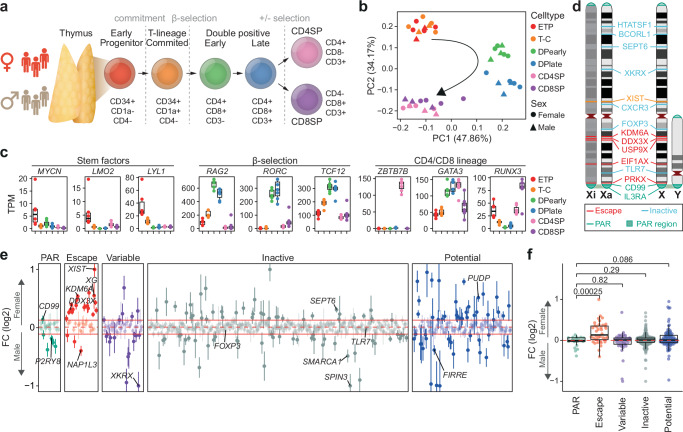
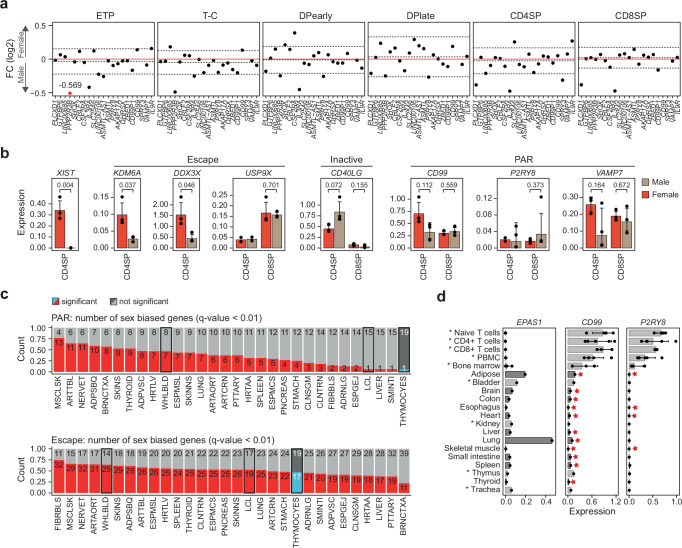
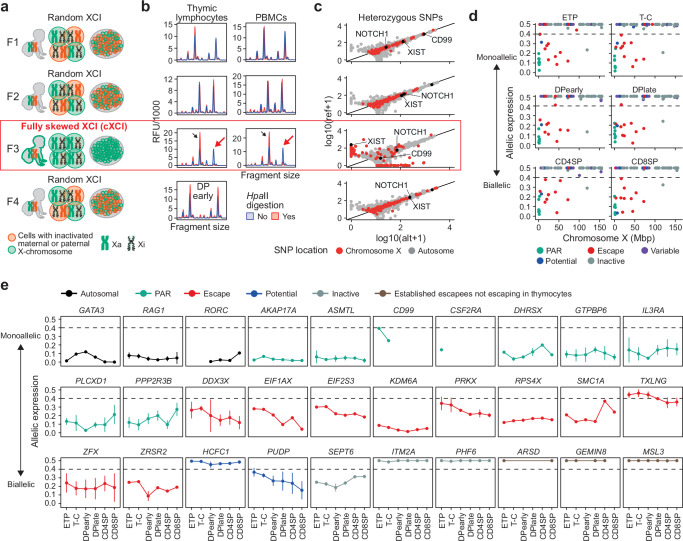
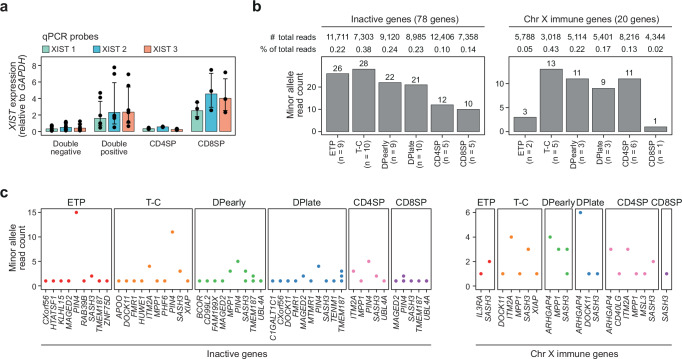
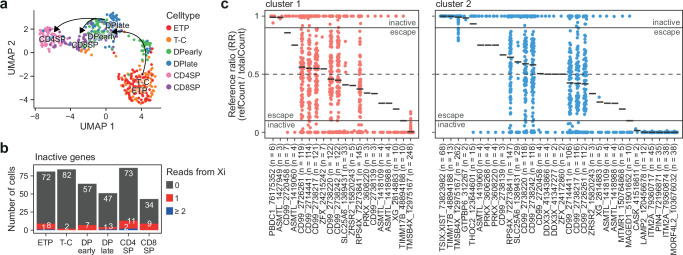
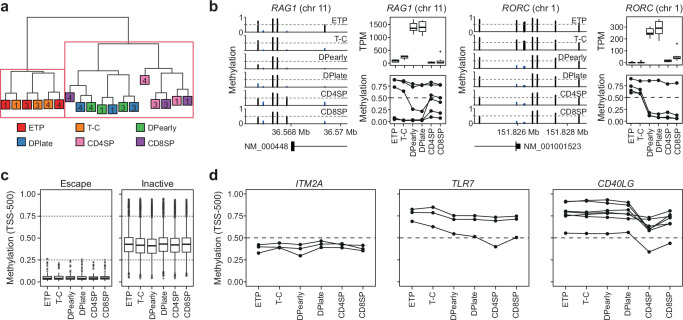
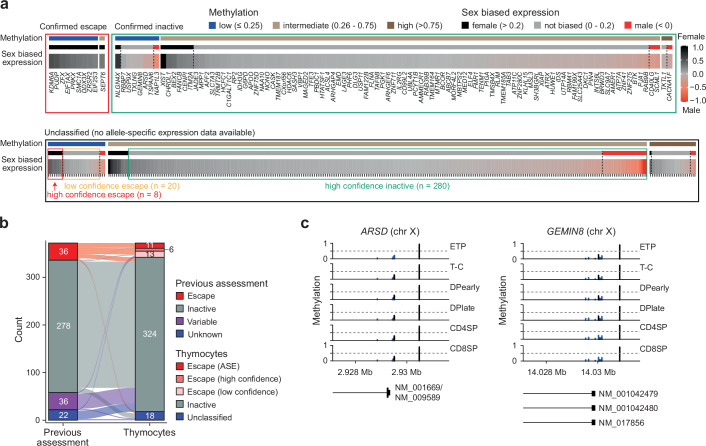
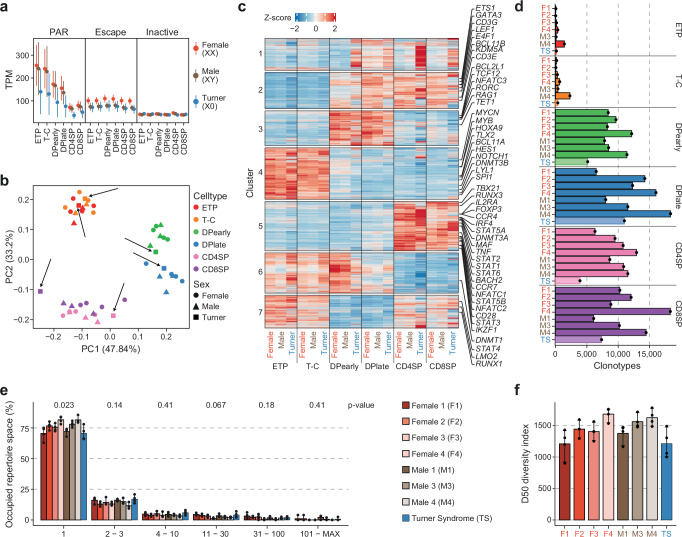
Females exhibit a more robust immune response to both self-antigens and non-self-antigens than males, resulting in a higher prevalence of autoimmune diseases but more effective responses against infection. Increased expression of X-linked immune genes in female T cells is thought to underlie this enhanced response. Here we isolate thymocytes from pediatric thymi of healthy males (46, XY), females (46, XX), a female with completely skewed X-chromosome inactivation (46, XX, cXCI) and a female with Turner syndrome (45, X0). Using whole exome sequencing, RNA sequencing and DNA methylation data, we present a sex-aware expression profile of T cell development and generate a high-resolution map of escape from X-chromosome inactivation (XCI). Unexpectedly, XCI is transcriptionally and epigenetically stable throughout T cell development, and is independent of expression of XIST, the lncRNA responsible for XCI initiation during early embryonic development. In thymocytes, several genes known to escape XCI are expressed from only one X-chromosome. Additionally, we further reveal that a second X-chromosome is dispensable for T cell development. Our study thus provides a high-resolution map of XCI during human development and suggests a re-evaluation of XCI in sex differences in T cell function.
© 2024. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
