

Tracking transcription-translation coupling in real time
- PMID: 39633055
- PMCID: PMC11711091
- DOI: 10.1038/s41586-024-08308-w
Tracking transcription-translation coupling in real time
Abstract
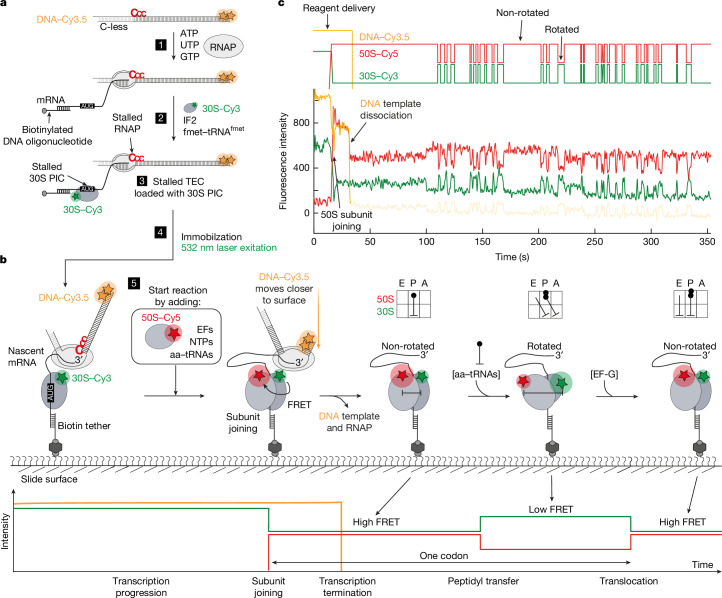
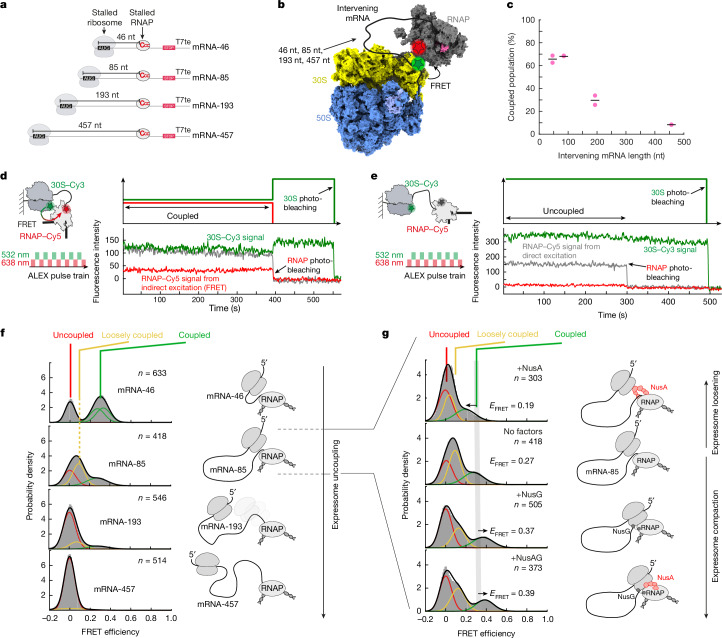
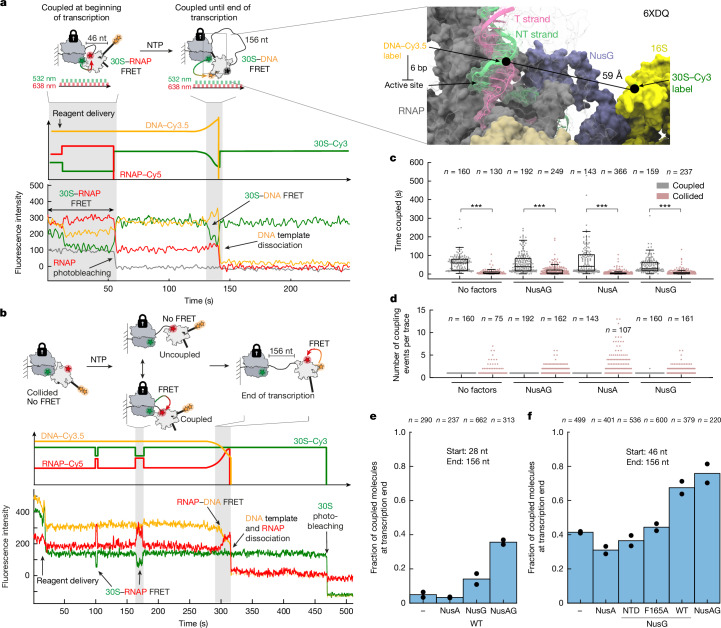
A central question in biology is how macromolecular machines function cooperatively. In bacteria, transcription and translation occur in the same cellular compartment, and can be physically and functionally coupled1-4. Although high-resolution structures of the ribosome-RNA polymerase (RNAP) complex have provided initial mechanistic insights into the coupling process5-10, we lack knowledge of how these structural snapshots are placed along a dynamic reaction trajectory. Here we reconstitute a complete and active transcription-translation system and develop multi-colour single-molecule fluorescence microscopy experiments to directly and simultaneously track transcription elongation, translation elongation and the physical and functional coupling between the ribosome and the RNAP in real time. Our data show that physical coupling between ribosome and RNAP can occur over hundreds of nucleotides of intervening mRNA by mRNA looping, a process facilitated by NusG. We detect active transcription elongation during mRNA looping and show that NusA-paused RNAPs can be activated by the ribosome by long-range physical coupling. Conversely, the ribosome slows down while colliding with the RNAP. We hereby provide an alternative explanation for how the ribosome can efficiently rescue RNAP from frequent pausing without requiring collisions by a closely trailing ribosome. Overall, our dynamic data mechanistically highlight an example of how two central macromolecular machineries, the ribosome and RNAP, can physically and functionally cooperate to optimize gene expression.
© 2024. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures












References
-
- Miller, O. L., Hamkalo, B. A. & Thomas, C. A. Visualization of bacterial genes in action. Science169, 392–395 (1970). - PubMed
-
- Blaha, G. M. & Wade, J. T. Transcription–translation coupling in bacteria. Annu. Rev. Genet.56, 187–205 (2022). - PubMed
-
- Burmann, B. M. et al. A NusE:NusG complex links transcription and translation. Science328, 501–504 (2010). - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
