

Revisiting mutational resistance to ampicillin and cefotaxime in Haemophilus influenzae
- PMID: 39633433
- PMCID: PMC11616347
- DOI: 10.1186/s13073-024-01406-4
Revisiting mutational resistance to ampicillin and cefotaxime in Haemophilus influenzae
Abstract
Background: Haemophilus influenzae is an opportunistic bacterial pathogen that can cause severe respiratory tract and invasive infections. The emergence of β-lactamase-negative ampicillin-resistant (BLNAR) strains and unclear correlations between genotypic (i.e., gBLNAR) and phenotypic resistance are challenging empirical treatments and patient management. Thus, we sought to revisit molecular resistance mechanisms and to identify new resistance determinants of H. influenzae.
Methods: We performed a systematic meta-analysis of H. influenzae isolates (n = 291) to quantify the association of phenotypic ampicillin and cefotaxime resistance with previously defined resistance groups, i.e., specific substitution patterns of the penicillin binding protein PBP3, encoded by ftsI. Using phylogenomics and a genome-wide association study (GWAS), we investigated evolutionary trajectories and novel resistance determinants in a public global cohort (n = 555) and a new clinical cohort from three European centers (n = 298), respectively.
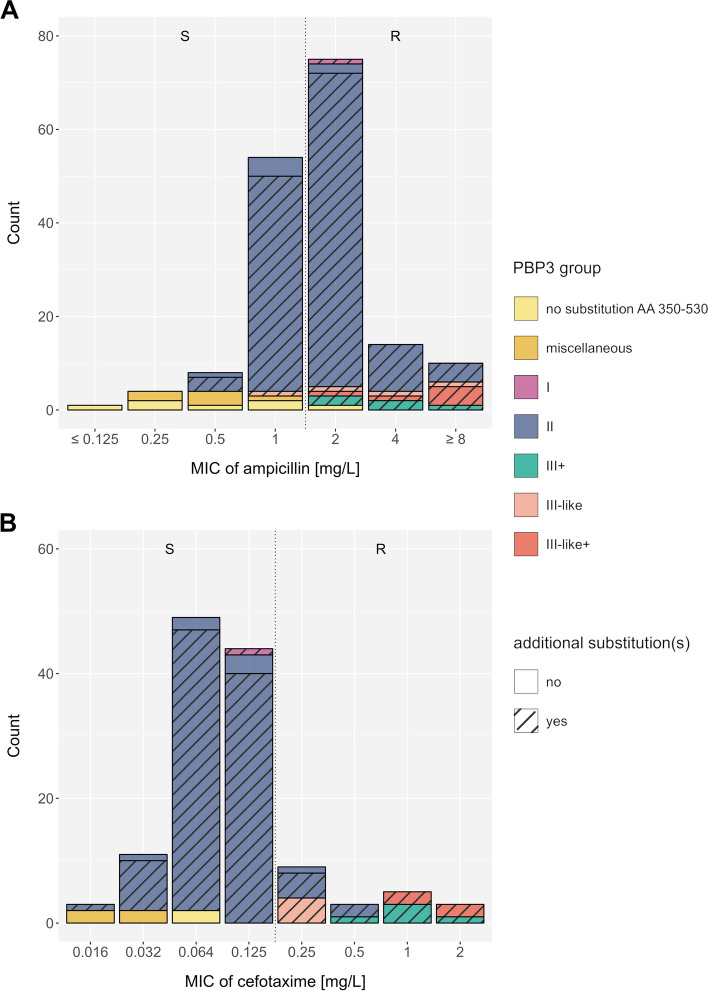
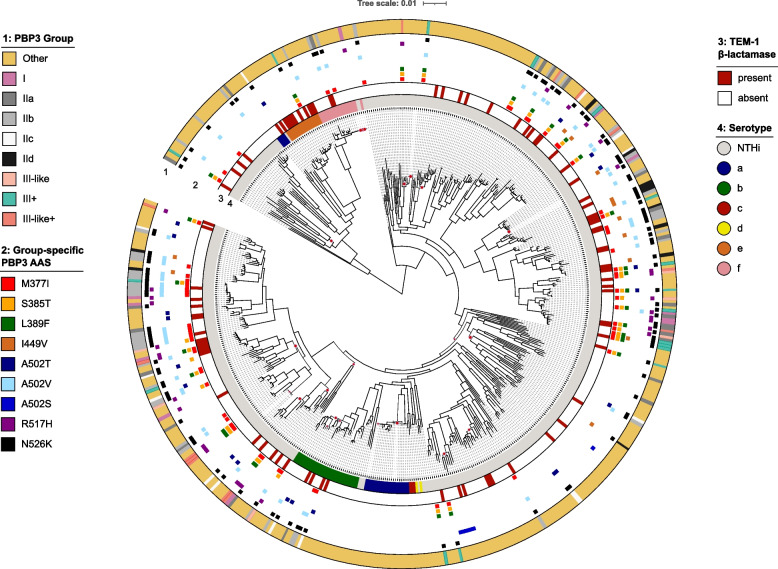
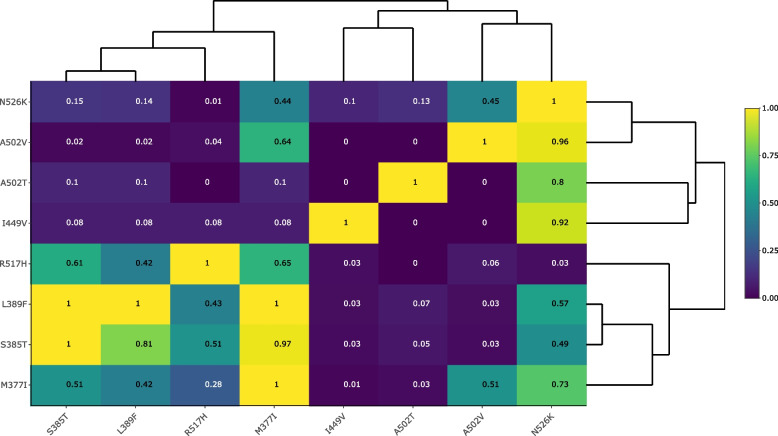
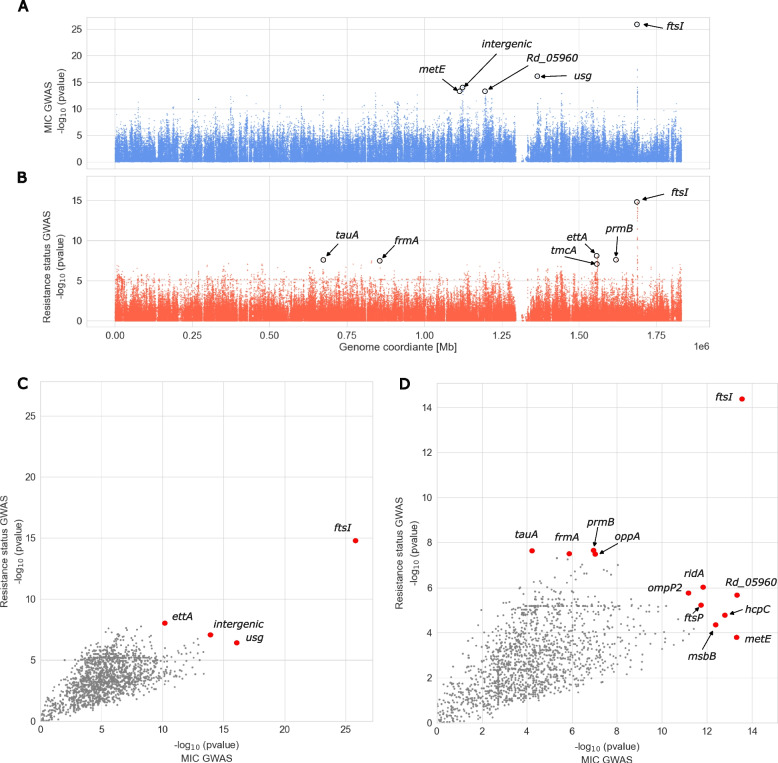
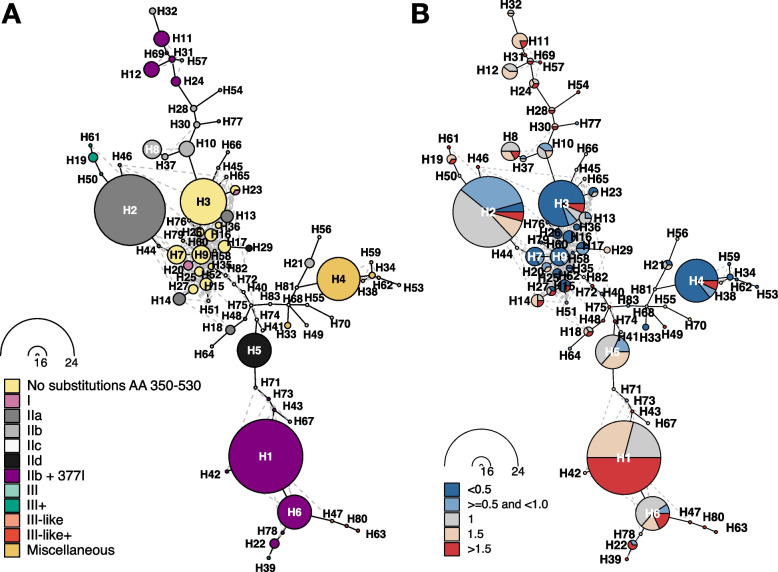
Results: Our meta-analysis confirmed that PBP3 group II- and group III-related isolates were significantly associated with phenotypic resistance to ampicillin (p < 0.001), while only group III-related isolates were associated with resistance to cefotaxime (p = 0.02). The vast majority of H. influenzae isolates not classified into a PBP3 resistance group were ampicillin and cefotaxime susceptible. However, particularly group II isolates had low specificities (< 16%) to rule in ampicillin resistance due to clinical breakpoints classifying many of them as phenotypically susceptible. We found indications for positive selection of multiple PBP3 substitutions, which evolved independently and often step-wise in different phylogenetic clades. Beyond ftsI, other possible candidate genes (e.g., oppA, ridA, and ompP2) were moderately associated with ampicillin resistance in the GWAS. The PBP3 substitutions M377I, A502V, N526K, V547I, and N569S were most strongly related to ampicillin resistance and occurred in combination in the most prevalent resistant haplotype H1 in our clinical cohort.
Conclusions: Gradient agar diffusion strips and broth microdilution assays do not consistently classify isolates from PBP3 groups as phenotypically resistant. Consequently, when the minimum inhibitory concentration is close to the clinical breakpoints, and genotypic data is available, PBP3 resistance groups should be prioritized over susceptible phenotypic results for ampicillin. The implications on treatment outcome and bacterial fitness of other extended PBP3 substitution patterns and novel candidate genes need to be determined.
Keywords: Haemophilus influenzae; Ampicillin resistance; BLNAR; Cefotaxime resistance; Genome-wide association study; Haplotype networks; Literature review; MIC; PBP3; Phylogenomics; Resistance groups.
© 2024. The Author(s).
Conflict of interest statement
Declarations. Ethics approval and consent to participate: The need for an ethics votum was waived by the ethics committee of the University of Wuerzburg (ref. 20240625 01) for the retrospective study of Haemophilus sp. isolated during routine clinical diagnostics (anonymized biomaterials). Therefore, the study did not require direct informed consent. All data were obtained in a blinded manner. The Lübeck study protocol was in addition approved by the local ethics committee of the University of Lübeck (Az 13-072). The research was conducted in accordance with the Declaration of Helsinki. Consent for publication: Not applicable. Competing interests: The authors declare that they do not have any competing interests.
Figures
References
-
- Factsheet about Invasive Haemophilus influenzae disease. 2017. Available from: https://www.ecdc.europa.eu/en/invasive-haemophilus-influenzae-disease/facts. Cited 6 Dec 2023.
-
- Kiedrowska M, Foryś WJ, Gołębiewska A, Waśko I, Ronkiewicz P, Kuch A, et al. Antimicrobial resistance among Haemophilus influenzae isolates responsible for lower respiratory tract infections in Poland, 2005–2019. Eur J Clin Microbiol Infect Dis Off Publ Eur Soc Clin Microbiol. 2022;41:961–9. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
