

Acute stress activates basolateral amygdala neurons expressing corticotropin-releasing hormone receptor type 1 (CRHR1): Topographical distribution and projection-specific activation in male and female rats
- PMID: 39634490
- PMCID: PMC11615582
- DOI: 10.1016/j.ynstr.2024.100694
Acute stress activates basolateral amygdala neurons expressing corticotropin-releasing hormone receptor type 1 (CRHR1): Topographical distribution and projection-specific activation in male and female rats
Abstract
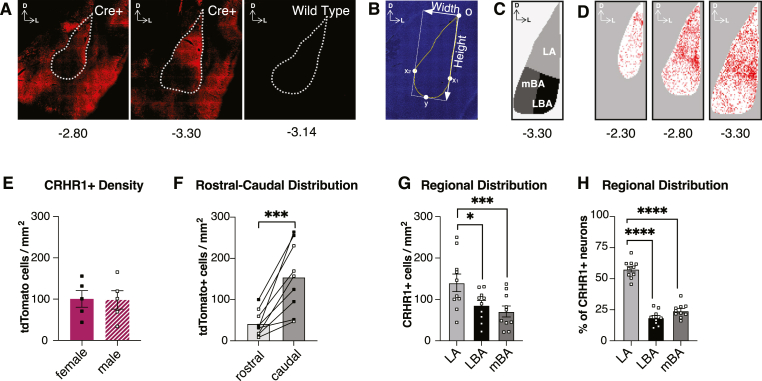
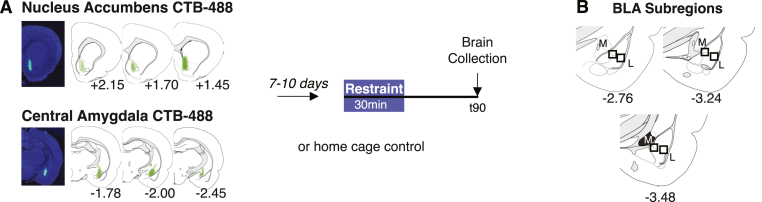
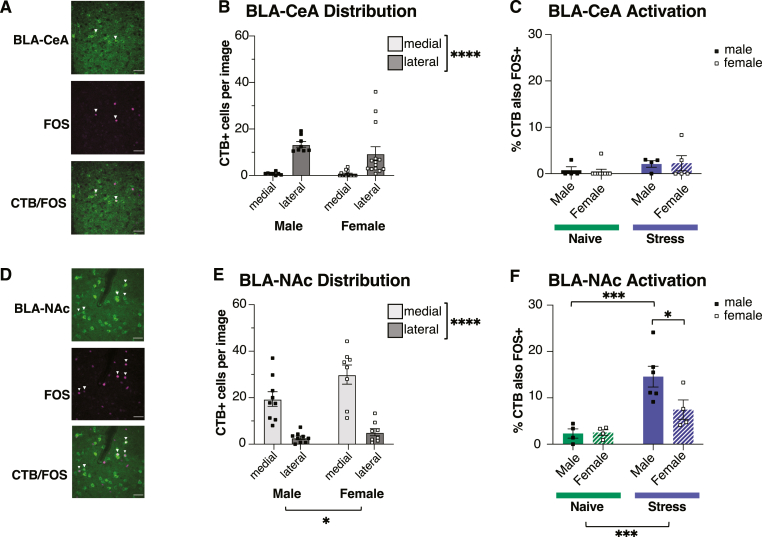
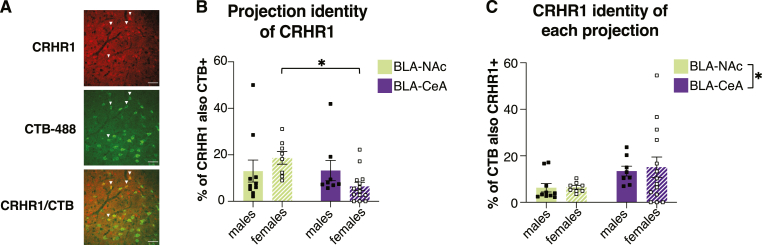
Although the basolateral amygdala (BLA) and corticotropin releasing hormone receptor type I (CRHR1) signaling are both central to the stress response, the spatial and circuit-specific distribution of CRHR1 have not been identified in the BLA at a high resolution. We used transgenic male and female CRHR1-Cre-tdTomato rats to topographically map the distribution of BLACRHR1 neurons and identify whether they are activated by acute stress. Additionally, we used the BLA circuits projecting to the central amygdala (CeA) and nucleus accumbens (NAc) as a model to test circuit-specific expression of CRHR1 in the BLA. We established several key findings. First, CRHR1 had the strongest expression in the lateral amygdala and in caudal portions of the BLA. Second, acute restraint stress increased FOS expression of CRHR1 neurons, and stress-induced activation was particularly strong in medial subregions of the BLA. Third, stress significantly increased FOS expression on BLA-NAc, but not BLA-CeA projectors, and BLA-NAc activation was more robust in males than females. Finally, CRHR1 was expressed on a subset of BLA-CeA and BLA-NAc projection neurons. Collectively, this expands our understanding of BLA molecular- and circuit-specific activation patterns following acute stress.
© 2024 The Authors. Published by Elsevier Inc.
Conflict of interest statement
Authors have no conflicts of interests to report.
Figures

Similar articles
-
Targeted overexpression of CRH receptor subtype 1 in central amygdala neurons: effect on alcohol-seeking behavior.Psychopharmacology (Berl). 2018 Jun;235(6):1821-1833. doi: 10.1007/s00213-018-4908-6. Epub 2018 Apr 26. Psychopharmacology (Berl). 2018. PMID: 29700576 Free PMC article.
-
Optogenetic Manipulations of Amygdala Neurons Modulate Spinal Nociceptive Processing and Behavior Under Normal Conditions and in an Arthritis Pain Model.Front Pharmacol. 2021 May 25;12:668337. doi: 10.3389/fphar.2021.668337. eCollection 2021. Front Pharmacol. 2021. PMID: 34113253 Free PMC article.
-
Activation of Basolateral Amygdala to Nucleus Accumbens Projection Neurons Attenuates Chronic Corticosterone-Induced Behavioral Deficits in Male Mice.Front Behav Neurosci. 2021 Feb 24;15:643272. doi: 10.3389/fnbeh.2021.643272. eCollection 2021. Front Behav Neurosci. 2021. PMID: 33716685 Free PMC article.
-
Generation of a CRF1-Cre transgenic rat and the role of central amygdala CRF1 cells in nociception and anxiety-like behavior.Elife. 2022 Apr 7;11:e67822. doi: 10.7554/eLife.67822. Elife. 2022. PMID: 35389341 Free PMC article.
-
New viral-genetic mapping uncovers an enrichment of corticotropin-releasing hormone-expressing neuronal inputs to the nucleus accumbens from stress-related brain regions.J Comp Neurol. 2019 Oct 15;527(15):2474-2487. doi: 10.1002/cne.24676. Epub 2019 Mar 30. J Comp Neurol. 2019. PMID: 30861133 Free PMC article.
References
-
- Agoglia A.E., Zhu M., Ying R., Sidhu H., Natividad L.A., Wolfe S.A., Buczynski M.W., Contet C., Parsons L.H., Roberto M., Herman M.A. Corticotropin-releasing factor receptor-1 neurons in the lateral amygdala display selective sensitivity to acute and chronic ethanol exposure. Eneuro. 2020;7(2) doi: 10.1523/ENEURO.0420-19.2020. ENEURO.0420-19.2020. - DOI - PMC - PubMed
-
- Atsak P., Hauer D., Campolongo P., Schelling G., Fornari R.V., Roozendaal B. Endocannabinoid signaling within the basolateral amygdala integrates multiple stress hormone effects on memory consolidation. Neuropsychopharmacology: Off. Publ. Am. Coll. Neuropsychopharmacol. 2015;40(6):1485–1494. doi: 10.1038/npp.2014.334. - DOI - PMC - PubMed
-
- Aukema R.J., Petrie G.N., Matarasso A.K., Baglot S.L., Molina L.A., Füzesi T., Kadhim S., Nastase A.S., Rodriguez Reyes I., Bains J.S., Morena M., Bruchas M.R., Hill M.N. Identification of a stress-responsive subregion of the basolateral amygdala in male rats. Neuropsychopharmacology: Off. Publ. Am. Coll. Neuropsychopharmacol. 2024 doi: 10.1038/s41386-024-01927-x. - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
